

INTRODUCTION
A fossil species is a hypothesis about the inclusiveness of a hypodigm defined by one or more fossil specimens (Simpson, 1940). A phylogenetic analysis is a formalised criterion for discriminating among alternative evolutionary frameworks all derived from the same set of hypotheses (Hennig, 1966). Since the operational taxonomic units included in phylogenetic analyses could be fossil species, the ways fossil hypodigms are defined are priors that shape (and might bias) the result of a phylogenetic analysis. From a phylogenetic perspective, the discussion on the alpha taxonomy of a fossil assemblage (variably polarised among “lumping” and “splitting” approaches; e.g., see discussion in Ibrahim et al., 2020a) represents an implicit (often unrecognised) statement on how the phylogenetic analysis is aprioristically defined. From this perspective, the taxonomic nomenclature followed by the authors is a distinct issue, albeit related: even under an “under-splitting” approach (sensu Ibrahim et al., 2020a) the data set could be “over-split” for methodological purposes (e.g., using operational units at the individual level of biological organisation, e.g., Cau, 2017). Distinct from the alpha taxonomy followed, the “methodological lumping prior” of a phylogenetic analysis is the amount of assumptions in assembling the taxon sample from the fossil data. Although the impact of alternative splitting approaches on the alpha taxonomy of a sample has been discussed (e.g., Ibrahim et al., 2020a), the effects of the alternative methodological lumping priors on the results of a phylogenetic analysis have received less attention (e.g., Cau, 2024; Kellermann et al., 2025). Here, we test the impact of methodological lumping priors on part of the fauna which was the object of the taxonomic discussion by Ibrahim et al. (2020a), the predatory dinosaur communities from the “mid-Cretaceous” of northern Africa (Stromer, 1934; Sereno et al., 1996; Russell, 1996; Sereno & Brusatte, 2008; Dyke, 2010; Chiarenza & Cau, 2016; Ibrahim et al., 2020a).
MATERIALS AND METHODS
We describe new fossil dinosaur material collected from the Cretaceous of the Errachidia Province, Morocco (Figs. 4-7). The material includes unassociated cranial and postcranial bones belonging to large-bodied theropods. All the material was collected in the “Kem Kem beds” (sensu Ibrahim et al., 2020a) by local collectors, acquired in the form of unprepared material from Moroccan fossil dealers and prepared by one of us (AP). The material was collected southeast of Taouz in the area known by the locals as “the red site”. With the exclusion of the lithology of the associated matrix removed during preparation (see below), we have no additional information regarding the exact stratigraphic position of the fossil material (see Ibrahim et al., 2020a), and no evidence about the association of multiple elements in cases when size and presence of apomorphic traits could support such attribution. All the material was exported according to the Moroccan and European legislation, and subsequently prepared at Ophis Museo Paleontologico e Centro Erpetologico in Teramo (Italy).
Geological nomenclature
The stratigraphic nomenclature of the “mid-Cretaceous” fossil-bearing units from the Moroccan-Algerian border is controversial and still debated (see historical reviews in Cavin et al., 2010; and Ibrahim et al., 2020a). This problem is exacerbated by the lack of accurate stratigraphic information associated with the majority of the surface-collected material from the “Kem Kem beds” (Dyke, 2010). Furthermore, the same authors used alternative and challenging stratigraphic zonations and nomenclature in subsequent publications, implicitly addressing the persistence of this problem (e.g., compare the stratigraphic nomenclature proposed by Ibrahim et al., 2020a, with the alternative nomenclature followed by Smith et al., 2023). Pending a consensus on the Kem Kem stratigraphy, the referral of the surface-sampled material from the various “mid-Cretaceous” localities in Morocco to a particular zonation system is in most cases arbitrary and non-testable. Although Cavin et al. (2010) explicitly recognised a compound nature for the “Kem Kem Vertebrate Assemblage” (sensu Ibrahim et al., 2020a), later studies were unable to recognise any faunal partition among the sampled localities (Ibrahim et al., 2020a). We thus refer the described fossil material to the “Kem Kem Assemblage” (a consensus between Cavin et al., 2010, and Ibrahim et al., 2020a), a term which focuses on the vertebrate fossil material collected from the “Kem Kem beds” but does not make any explicit reference to a particular stratigraphic framework. This approach addresses the potential time-averaged mixture of subsequent discrete faunas represented by the described fossil material (Cavin et al., 2010), but remains agnostic about the general zonation framework (e.g., Ibrahim et al., 2020a, versus Smith et al., 2023) and about the – often unknown – particular stratigraphic placement of the individual specimens collected (Dyke, 2010). We encourage the use of this term in cases, like those studied here, of fossil material lacking accurate stratigraphic data (due to a lack of collecting information and/or absence of consensus on the zonation system). This terminology does not constitute an alternative to any of the discussed stratigraphic frameworks, yet it makes it explicit the problematic stratigraphy affecting a significant part of the “Kem Kem” fossils reported in literature.
Phylogenetic Nomenclature
We introduce a terminology differentiating among the inclusiveness of the operational taxonomic units.
Operational Taxonomic Sub-Unit (OTSU). Any Operational Taxonomic Unit (OTU) based on a subset of the scores of another OTU. Example: an OTU coded uniquely from the skull material of Allosaurus (and leaving all non-skull scores as “unknown”) is an OTSU of the Allosaurus OTU.
Non-Overlapping Operational Taxonomic Sub-Units (NOTSUs). Two or more OTSUs based on the same OTU, which do not overlap in any coded character. Example: the Allosaurus OTSU coded uniquely from the forelimb material and the Allosaurus OTSU coded uniquely from the hindlimb material are both NOTSUs of Allosaurus OTU.
Complementary Operational Taxonomic Sub-Unit (COTSU). Two or more NOTSUs which combined produce the complete score of the original OTU. Example: assuming a character list of 200 character statements, the Allosaurus NOTSU based on the character scores 1-100 and the Allosaurus NOTSU based on characters scores 101-200 are COTSUs of the Allosaurus OTU.
Taxon sample
In order to test the impact of alternative assumptions on the alpha taxonomy of the mid-Cretaceous theropod communities from northern Africa, we used a recently-published data set focusing on non-neornithine theropod relationships (Cau, 2024). Both the taxon sample in the original data set and that used here (i.e., 195 taxonomic units vs 3888 characters, see Supplementary Material) have been defined in order to minimise a priori assumptions on the monophyly and inclusiveness of the included taxa, and follow a coding protocol which prevents topological artifacts due to ontogeny-biased miscoding (see details in Cau, 2024). As in the original analysis, several taxa from the “mid-Cretaceous” of northern Africa were split into distinct OTUs to prevent a priori introduction in the sample of “chimaeras” (see supplementary information in Cau, 2024). A similar approach was independently followed by Kellermann et al. (2025). Yet, we disagree with the latter in the inclusiveness of Eocarcharia and Megaraptor. The former one will be discuss in detail, below. The two distinct taxonomic units of Megaraptor used by Kellermann et al. (2025) are here considered mature and immature semaphoronts of the same taxon and coded in the relative partitions of the same taxonomic unit (as in Cau, 2024, in agreement with Porfiri et al., 2014). We have reviewed the ontogenetic stage of some specimens included in the original data set (Cau, 2024). The holotype of Garudimimus shows all braincase elements disarticulated, a partially-fused sacral series and most of the presacral neural arches unfused to the centra, supporting a juvenile status (Kobayashi & Barsbold, 2006). Following Hendrickx et al. (2024), Noasaurus holotype is considered an immature individual. Accordingly, both Garudimimus and Noasaurus have been recoded in the immature partition of the data matrix. The character codings of the Egyptian specimen of Carcharodontosaurus (sensu Stromer, 1931; renamed as Tameryraptor by Kellermann et al., 2025), Shaochilong and Labocania were updated following Kellermann et al. (2025) and Rivera-Sylva & Longrich (2024). As discussed below, we consider the two frontoparietal morphotypes from the Kem Kem unit described by Arden et al. (2019) as different semaphoronts of “bone taxon A” first described by Russell (1996). Although Ibrahim et al. (2020a) questioned their theropod affinities, no evidence for a crocodylomorph status was provided by those authors. We follow Arden et al. (2019) and Lacerda et al. (2024) and included an OTU based on “bone taxon A” morphs in the data matrix.
The status of both Bahariasaurus and Deltadromeus is particularly controversial (see reviews in Carrano & Sampson, 2008; Chiarenza & Cau, 2016; Ibrahim et al., 2020a). Stromer (1934) described a series of associated vertebrae from the Bahariya Formation which includes posterior dorsal vertebrae showing the diagnostic morphology of Bahariasaurus type material (specimen SNSB-BSPG1912VIII62, Stromer, 1934), some caudal vertebrae bearing pleurocoels, and an enigmatic neural arch illustrated in posterior view (see Fig. 16 in plate II in Stromer, 1934). The latter closely matches the morphology of the theropod cervicodorsal elements (e.g., O’Connor, 2007; Aranciaga Rolando et al., 2021), and is thus referred to that region of the axial column. We refer the above mentioned series of associated bones to Bahariasaurus. Following Ibrahim et al. (2020a), the holotype of Deltadromeus agilis is re-interpreted as immature: accordingly, the code string of the type material used in Cau (2024) has been recoded in the immature partition of the data matrix. Furthermore, we agree with Ibrahim et al. (2020a) that part of the material referred to Bahariasaurus by Stromer (1934) broadly overlaps in morphology with Deltadromeus holotype. The differences between the two hypodigms (e.g., Cau & Chiarenza, 2016) are compatible with those observed among other theropod populations (e.g., Griffin, 2018). Accordingly, the scapula, coracoid and associated caudal vertebrae (specimen SNSB-BSPG1912VIII60, Stromer, 1934), the femur (specimen SNSB-BSPG1912VIII69, Stromer, 1934) and the fibula (specimen SNSB-BSPG1912VIII70, Stromer, 1934) are considered mature specimens of the same taxon including the Deltadromeus type material in its immature partition. Furthermore, the exceptionally long and gracile humerus (i.e., SNSB-BSPG1912VIII177) described by Stromer (1934) very closely recalls the humerus of Deltadromeus (Ibrahim et al., 2020a) and it thus referred to a mature individual of the same taxon. Yet, we follow Carrano & Sampson (2008) and Apesteguia et al. (2016) and disagree with Sereno et al. (1996) and Ibrahim et al. (2020a) in the interpretation of the distal end of the Deltadromeus holotypic “pubis”, which is instead considered the distal end of the ischium. The true pubic foot of Deltadromeus holotype has been unambiguously identified by the associated gastralia (Apesteguia et al., 2016).
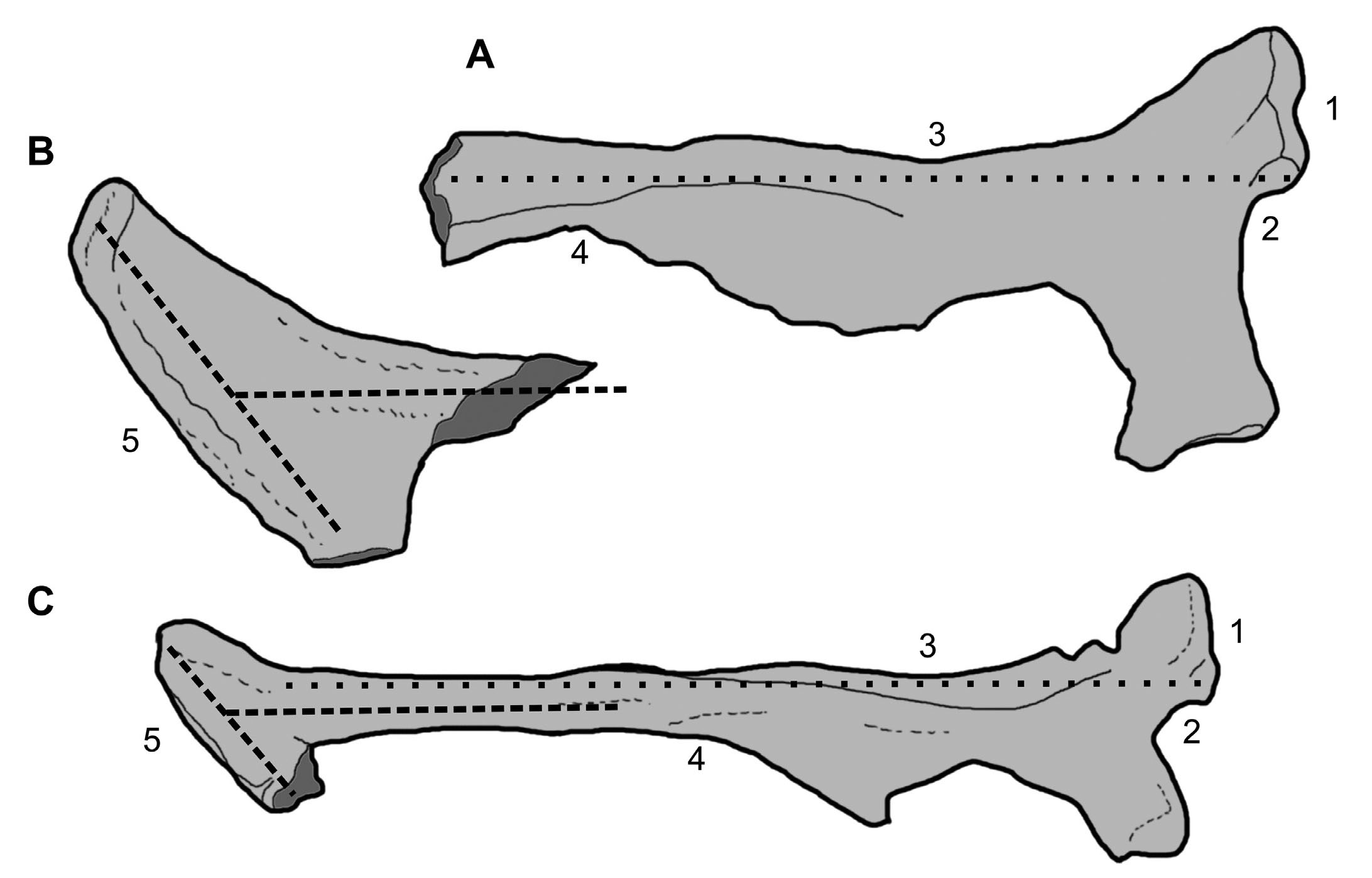
Here, we show that the revised interpretation of Deltadromeus ischium (Carrano & Sampson, 2008) provides the basis for resolving its status relative to Bahariasaurus. Although the two holotypes of Bahariasaurus and Deltadromeus directly overlap only in portions of the pelvic bones (Carrano & Sampson, 2008; Apesteguia et al., 2016), the ischial foot of Deltadromeus closely matches the distal end of a theropod bone from the Bahariya Formation, i.e. specimen SNSB-BSPG1912VIII82, erroneously interpreted as a pubis by Stromer (1934) and which instead we consider an ischium. Ischial features in specimen SNSB-BSPG1912VIII82 include the presence of the obturator process, the concave acetabular margin, the very narrow and proximally extended apron, and the very close correspondence of its proximal end with the autapomorphic morphology of Bahariasaurus type ischium (Fig. 1; see also plate II in Stromer, 1934). Thus, SNSB-BSPG1912VIII82 shows: 1) the peculiar features of Bahariasaurus holotype’s ischium in its proximal end (note that the latter bone lacked its distal end; Stromer, 1934), and 2) the peculiar features of Deltadromeus holotype’s ischium in its distal end (note that the proximal end of the latter bone is poorly preserved; Carrano & Sampson, 2008, contra Sereno et al., 1996). The specimen SNSB-BSPG1912VIII82 supports the referral of Bahariasaurus and Deltadromeus to the same taxon (Fig. 1), which is named Bahariasaurus by priority. Accordingly, we have included in the hypodigm of Bahariasaurus: the holotype of Deltadromeus, the material referred to Deltadromeus by Sereno et al. (1996) and Ibrahim et al. (2020a), and all theropod material from the Bahariya Formation described by Stromer (1934) which shares the derived features of at least one of the two holotypes (Fig. 2).
- Deltadromeus is a junior synonym of Bahariasaurus. (A) Preserved proximal end of the right ischium of Bahariasaurus ingens holotype BSP 1922X47 in lateral view (redrawn from Stromer, 1934). (B) Preserved distal end of the right ischium of Deltadromeus agilis holotype SGM-DIN 2 in lateral view (redrawn from Sereno et al., 1996). (C) Right ischium from the Bahariya Formation, BSP 1912VIII82 (originally considered the pubis of an indeterminate taxon by Stromer, 1934) in lateral view (redrawn from Stromer, 1934). The combination of features 1 to 5 supports the referral of the three ischia to the same taxon. 1: iliac peduncle anteroposteriorly twice longer than proximodistally deep; 2: acetabular margin shallow and angular at level of iliac peduncle; 3: posterior margin of shaft aligned with acetabular margin of iliac peduncle; 4: triangular obturator process distally confluent with ischial shaft without a distinct notch; 5: anteroposteriorly enlarged distal foot with long axis forming a 120° angle with the long axis of the ischial shaft. Figures A to C not at same scale. Broken bone margins in dark grey.
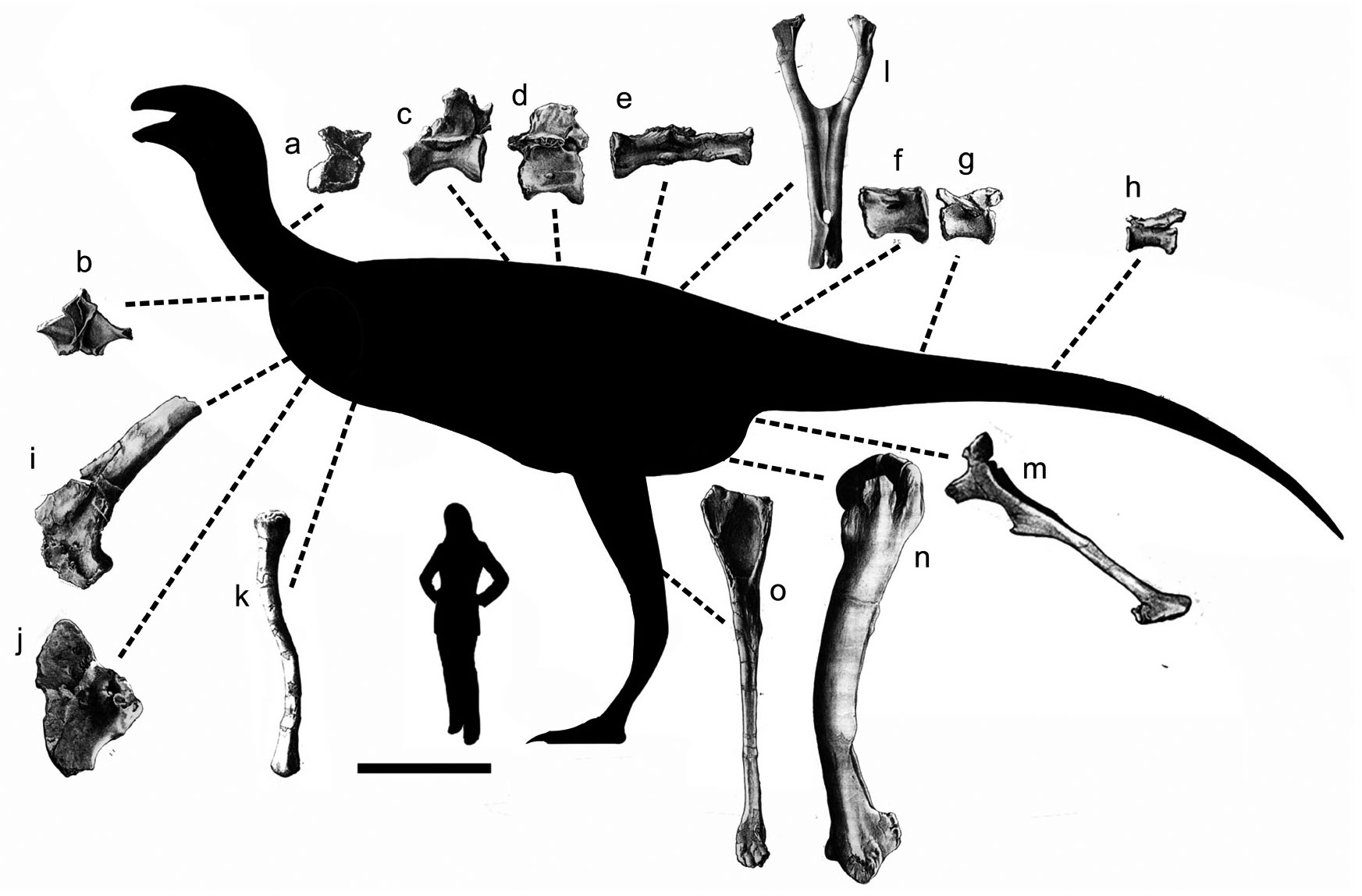
- Theropod bones from the Bahariya Formation included in Bahariasaurus ingens hypodigm and used in the phylogenetic analysis. Bahariasaurus silhouette based on the largest femur collected (Stromer, 1934). Scale bar = 1 m. (a): posterior cervical vertebra in left lateral view; (b): cervicodorsal neural arch in posterior view; (c) and (d): dorsal vertebrae in left lateral view; (e): partial sacrum in ventral view; (f)-(h): caudal vertebrae in lateral view; (i) partial left scapula in lateral view; (j) left coracoid in lateral view; (k) humerus in posterior view; (l): pubis in posterior view; (m): ischium in lateral view; (n): femur in lateral view; (o): fibula in medial view. (All pictures modified from Stromer, 1934; not at same scale)
We have also reviewed the status of two Jehol Biota theropods included in the taxon sample of Cau (2024). We consider Sinotyrannus (Ji et al., 2009) a junior synonym of Huaxiagnathus (Hwang et al., 2004). The hypodigm of Huaxiagnathus is based on immature “compsognathid-grade” specimens (Cau, 2024), whereas that of Sinotyrannus includes only a mature individual (Hwang et al., 2004; Ji et al., 2009). Among early-diverging coelurosaurs, Sinotyrannus shares with Huaxiagnathus the peculiar morphology of the ilium, which is dorsally convex, with a relatively short preacetabular process lacking the anteroventral hook (Hwang et al., 2004; Ji et al., 2009). Most of the differences between the two taxa are size-related or ontogenetically-controlled features, including the proportions of the premaxilla and maxilla, and the size and position of the maxillary fenestra, all matching the ontogenetic-controlled diversity expected for a tyrannosauroid growth series (e.g., Tyrannosaurus, Carr & Williamson, 2004). We thus consider “Sinotyrannus” as a mature morphotype of Huaxiagnathus: accordingly, the code string of the former has been included in the mature partition of the latter OTU.
Following Cau (2024), the species-level OTUs of some African theropods (i.e., Carcharodontosaurus saharicus, C. iguidensis, Eocarcharia dinops, Kryptops palaios, Spinosaurus aegyptiacus sensu Ibrahim et al., 2014) are replaced by a series of OTSUs: each OTSU has been coded uniquely from one specimen among those referred in literature to the relative species-level OTU. In some cases, the different OTSUs coded from the same taxon partially overlap (e.g., the OTU including the Spinosaurus neotype FSACKK11888, Ibrahim et al., 2014; 2020b; and that coded from Spinosaurus holotype, Stromer, 1915), because they share a series of scores relative to the presacral vertebrae. Yet, we provisionally kept them separate OTUs to test the hypothesis that they do not pertain to the same species (Evers et al., 2015). In other cases, the different OTSUs do not overlap in any score, and constitute the COTSUs of the original OTU (e.g., the two OTUs based on, respectively, the postorbital skull material including the type of Eocarcharia dinops and the non-associated maxilla referred to the same taxon; or the two OTUs based on, respectively, the holotype maxilla and the referred postcranium of Kryptops palaios; see Sereno & Brusatte, 2008, and Kellermann et al., 2025). Zitouni et al. (2019) referred a partial ilium from the Kem Kem units to Abelisauridae: following Ibrahim et al. (2020a) and Samathi (2025), this specimen is instead considered a spinosaurid and coded in the immature partition of the Kem Kem spinosaurine morphotype OTU also including Spinosaurus aegyptiacus neotype. The hypodigm of Saurophaganax maximus is chimaerical and includes sauropod and theropod bones collected from the same locality (Danison et al., 2024): accordingly, the taxon was removed from the analysis.
Several character statements used by Cau (2024) have been emended or replaced (see details in Supplementary Material).
The taxon sample was used to test the impact on topology of null hypotheses concerning the hypodigms of the mentioned African theropods. This “methodological splitting” approach did not enforce any constraint among the included OTUs, and thus evaluated the relationships among them with no assumptions on the number and inclusiveness of the taxa in the sample (Brusatte & Sereno, 2007; Sereno & Brusatte, 2008; Carrano et al., 2012; Ibrahim et al., 2014, 2020a; Paterna & Cau, 2022; Cau, 2024; Kellermann et al., 2025). The phylogenetic analysis was performed in TNT vers. 1.6 (Goloboff & Morales, 2023): the analysis performed 1000 “New Technology” runs followed by a second “Tree Bisection Reconnection” analysis exploring the tree sample produced by the first run. We sampled 10.000 shortest trees and used that sample to reconstruct a consensus topology. The most unstable OTUs included in the analysis were then pruned a posteriori from the saved trees: their affinities were evaluated relative to the reduced consensus topology based on the other OTUs.
In the Systematic Palaeontology section, we referred the specimens included in the phylogenetic analysis to the least inclusive clade containing all alternative placements of the specimen: in some cases (e.g., OPH2025), the referred clade was more inclusive than that inferred following a traditional apomorphy-based approach (i.e., positive overlap in size and morphology to material described in literature). Specimens too fragmentary for being included in the phylogenetic analysis were referred to a particular clade following the apomorphy-based approach. Specimens whose coding string resulted a redundant subset of another OTU(s) were excluded from the analysis (“safe taxonomic reduction” sensu Wilkinson, 1995).
We used the phylogenetic relationships among the sampled taxa as a proxy for calculating the similarity among the four best known “mid-Cretaceous” units from northern Africa (e.g., the Aptian-Albian Elhraz Formation from Niger, the Cenomanian Bahariya Formation from Egypt, the Cenomanian Eckhar Formation from Niger, and the Kem Kem Assemblage from the Aptian-Cenomanian of Morocco; Stromer, 1915, 1934; Russell, 1996; Sereno et al., 1996; Sereno et al., 2004; Brusatte & Sereno, 2007; Sereno & Brusatte, 2008). Unit similarity was calculated using the unweighted pair group method with arithmetic mean algorithm of the binary matrix expressing the phyletic similarity among the taxa sampled from each unit.
Institutional abbreviations
BMNH, Natural History Museum of London: Department of Palaeontology, UK; BSPG, Bayerische Staatssammlung fur Paläontologie und Geologie, Munchen, Germany; NMC, Canadian Museum of Nature, Ottawa, Canada (formerly NMC); DNM / DINO, Dinosaur National Monument, Jensen, Utah, USA; FSAC, Faculte des Sciences Aïn Chock, University Hassan II, Casablanca, Morocco; MCF-PVPH, Museo Municipal ‘Carmen Funes’, Paleontología de Vertebrados Plaza Huincul, Provincia de Neuquén, Argentina; ML, Museu da Lourinhã, Lourinhã, Portugal; MNN, Musée National de Boubou Hama, Niamey, Niger; MPEF-PV, Museo Paleontologico Egidio Feruglio, Trelew; MPM, Museo Paleontologico di Montevarchi, Italy; MSNM, Museo di Storia Naturale, Milano, Italy; MUCPV, Museo de la Universidad del Comahue, Argentina; NCSM, North Carolina State Museum of Natural Sciences, Raleigh, USA; OPH, Ophis Museo Paleontologico e Centro Erpetologico, Teramo, Italy; SGM, Ministére de l’Energie et des Mines, Rabat, Morocco; SMU, Southern Methodist University, Dallas; SNSB-BSPG, Bayerische Staatssammlung für Paläontologie und Historische Geologie, Munich, Germany; TMP, Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta, Canada; UCPC, University of Chicago Research Collection, Chicago, USA; UUVP, University of Utah, Vertebrate Paleontology Collection, Salt Lake City.
Other abbreviations
COTSU, Complementary Operational Taxonomic Sub-Unit; OTSU, Operational Taxonomic Sub-Unit; OTU, Operational Taxonomic Unit; NOTSU, Non-Overlapping Operational Taxonomic Sub-Unit.
RESULTS
The analysis reconstructed 10000 shortest trees of length 9397 steps each (tree length Consistency Index excluding uniformative characters, CI, = 0.2501; Retention Index, RI, = 0.5242) (Fig. 3).
- (A) Reduced strict consensus of the shortest trees found by the phylogenetic analysis. Letters indicate the pruned “wildcard” OTUs. (B) Similarity analysis of the four main “mid-Cretaceous” theropod communities from northern Africa based on the affinities among the sampled taxa. Numbers at nodes indicate the bootstrapping value (1000 replications). Alternative placements of the pruned OTUs marked by letters at branches; a: Angaturama; b: Carcharodontosaurus iguidensis (holotype maxilla); c: Eustreptospondylus; d: Fukuiraptor; e: Garudimimus; f: Irritator; g: Kem Kem spinosaurine rostrum morph (Dal Sasso et al., 2005); h: Noasaurus; i: Shidaisaurus; j: Taurovenator (referred specien MPCAPv8); k: Bone taxon A (Russell, 1996); l: Kem Kem cervical morph (Russell, 1996; this study); m: Compsognathus corallestris; n: Juravenator; o: OPH2025 (axis); p: OPH2120 (sacrum).
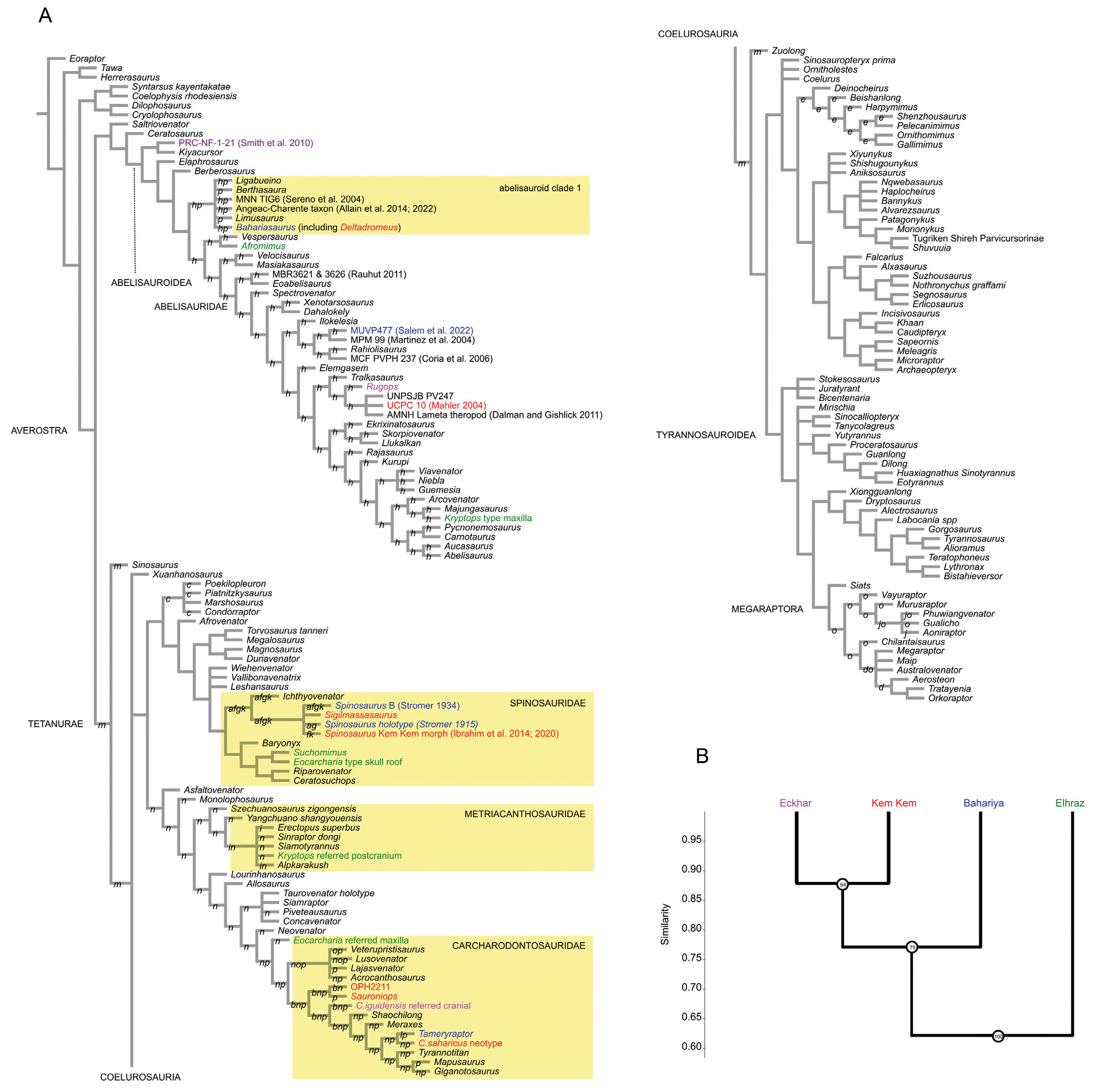
The phylogenetic relationships among the main theropod groups (i.e., Allosauroidea, Ceratosauria, Coelophysoidea, Coelurosauria, Megalosauroidea; Carrano & Sampson, 2008; Carrano et al., 2012) are in agreement with previous versions of this data set (e.g., Cau, 2024): the strict consensus of the shortest trees found reconstructed the same pectinate series, with coelophysoids sister taxon of averostrans among Neotheropoda, ceratosaurians sister group of tetanurans among Averostra, and coelurosaurs sister taxon of Carnosauria (the latter including allosauroids and megalosauroids) among tetanurans. The megaraptorans were reconstructed as a tyrannosauroid coelurosaurian clade, as in previous iterations of the data set (Cau, 2018; 2024). Differing from the previous result (Cau, 2024), the “bahariasaurids” (Stromer, 1934; Sereno et al., 1996; Apesteguia et al., 2016; Motta et al., 2016) were reconstructed as non-monophyletic, i.e., Bahariasaurus was placed among the early-diverging abelisauroids, whereas Aoniraptor and Gualicho were placed among Megaraptora. As in Cau (2024), the noasaurids sensu lato resulted a paraphyletic grade of Abelisauroidea forming a pectinate series leading to Abelisauridae. Yet, the very unstable placement of the fragmentary and immature Noasaurus OTU relative to other ceratosaurs in the trees sampled prevents fixation of the names Noasauridae and Noasaurinae to any of the branches of Abelisauroidea. We provisionally name the abelisauroid clade containing Bahariasaurus but excluding Masiakasaurus and abelisaurids as “abelisauroid clade 1”.
The hypodigm of Spinosaurus aegyptiacus (sensu lato, Ibrahim et al. 2014) was reconstructed as potentially monophyletic (i.e., all the northern African OTSUs clustered in the same terminal node) because it formed a soft polytomy with other non-African OTUs due to non-overlapping codes.
The two OTUs based on Carcharodontosaurus iguidensis hypodigm (Kellermann et al., 2025; see also the supplementary material in Cau, 2024) were reconstructed among Carcharodontosauridae, yet they did not cluster together neither with Carcharodontosaurus saharicus, in agreement with previous studies challenging Carcharodontosaurus monophyly (Chiarenza & Cau, 2016; Paterna & Cau, 2022; Kellermann et al., 2025).
Despite the revision of the Egyptian carcharodontosaur described by Stromer in 1931 (re-named as Tameryraptor by Kellermann et al., 2025), the latter resulted sister taxon of the Moroccan neotype of Carcharodontosaurus saharicus (Brusatte & Sereno, 2007) as reconstructed in Cau (2024), even following the revised codes suggested by Kellermann et al. (2025).
Both hypodigms of the Elrhaz Formation theropods Eocarcharia dinops and Kryptops palaios (Sereno & Brusatte, 2008) resulted chimaerical. As suggested by previous authors (e.g., Carrano et al., 2012; Kellermann et al., 2025), Kryptops palaios original hypodigm results based on an abelisaurid maxilla (the holotype) and allosauroid postcranial material. An original result of our analysis is the placement of the postcranial material referred to Kryptops to Metriacanthosauridae. Another novel result of our analysis is the placement of the skull roof of Eocarcharia (including the holotype postorbital) among the baryonychine spinosaurids, whereas the referred maxilla was confirmed among early-diverging carcharodontosaurids (Sereno & Brusatte, 2008).
Systematic palaeontology
Dinosauria Owen, 1842
Theropoda Marsh, 1881
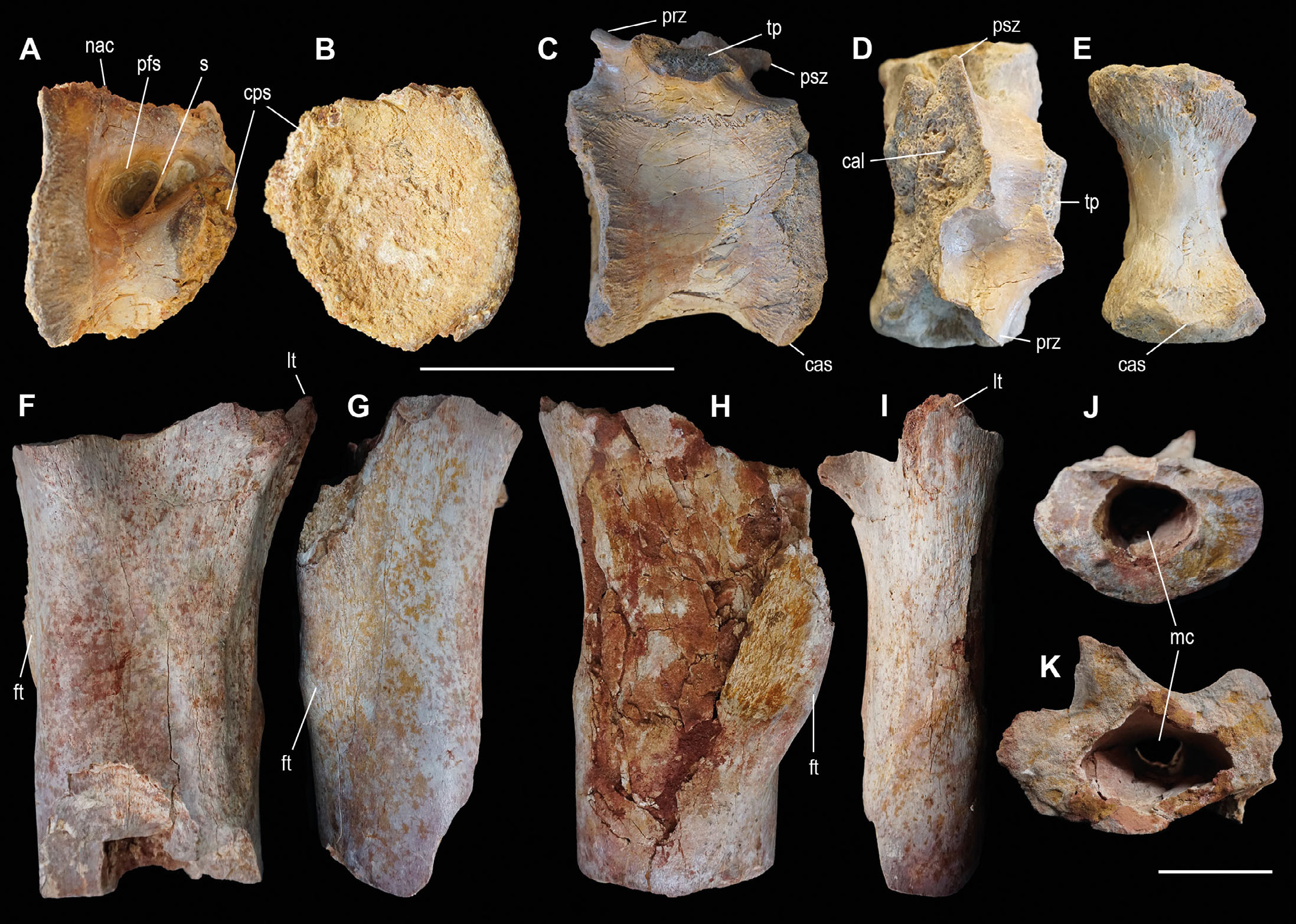
Quadratojugal. The specimen OPH2210 is a well-preserved right quadratojugal (Fig. 4A-B). It is flattened and mediolaterally compressed. The bone is concave medially, and convex laterally near the margin of the infratemporal fenestra. Laterally, the bone surface is overall smooth and pitted by microporosities. Small foramina and rugosities are scattered along the posteroventral corner of the bone. Anteriorly, just below the margin of the infratemporal fenestra, a descending diagonal line reveals the contact with the jugal, which extends to the posterior half of the bone’s ventral surface. Medially, most of the surface is smooth, except for the ventral and dorsal quadrate facets, and on the dorsal end of the squamosal process. At the level of the ventral facet for the quadrate, the bone reaches its maximum thickness. The margin of the quadrate-quadratojugal foramen is smooth and protrudes medially along the ventral border. The margin of the infratemporal fenestra is thin, further tapering to a sharp crest at the jugal process.
- Theropod material from the Kem Kem Assemblage. Right quadratojugal OPH2210 in lateral (A) and medial (B) view. Axis vertebra OPH2025 in right lateral (C) and anterior (B) view. Sacral vertebrae OPH2120 in lateral (E), ventral (F) and posterior (G) view. Left coracoid OPH2234 in lateral view (H). Left scapula OPH2233 in lateral view (I). Right femur OPH2107 in anterior (J) and lateral (K) view. Left metatarsal II OPH2213 in medial (L) and anterior (M) view. Scale bars = 100 mm. Abbreviations: ac, acromion; cf; coracoid foramen; cm, cranial margin; cps, camellate pneumatic structure; dp, diapophysis; dqc, dorsal quadratojugal contact; ft, fourth trochanter; gl, glenoid; ic, intercentrum; ifr, infratemporal fenestra rim; jc, jugal contact; lcd, lateral condyle; lt, lesser trochanter; mc, medullary cavity; mcd, medial condyle; nc, neural canal; nvf, neurovascular foramen; od, odontoid; pfs, pneumatic fossa; pp, parapophysis; qf, quadrate-quadratojugal foramen; s, septum; sp, squamosal process; vqc, ventral quadratojugal contact.
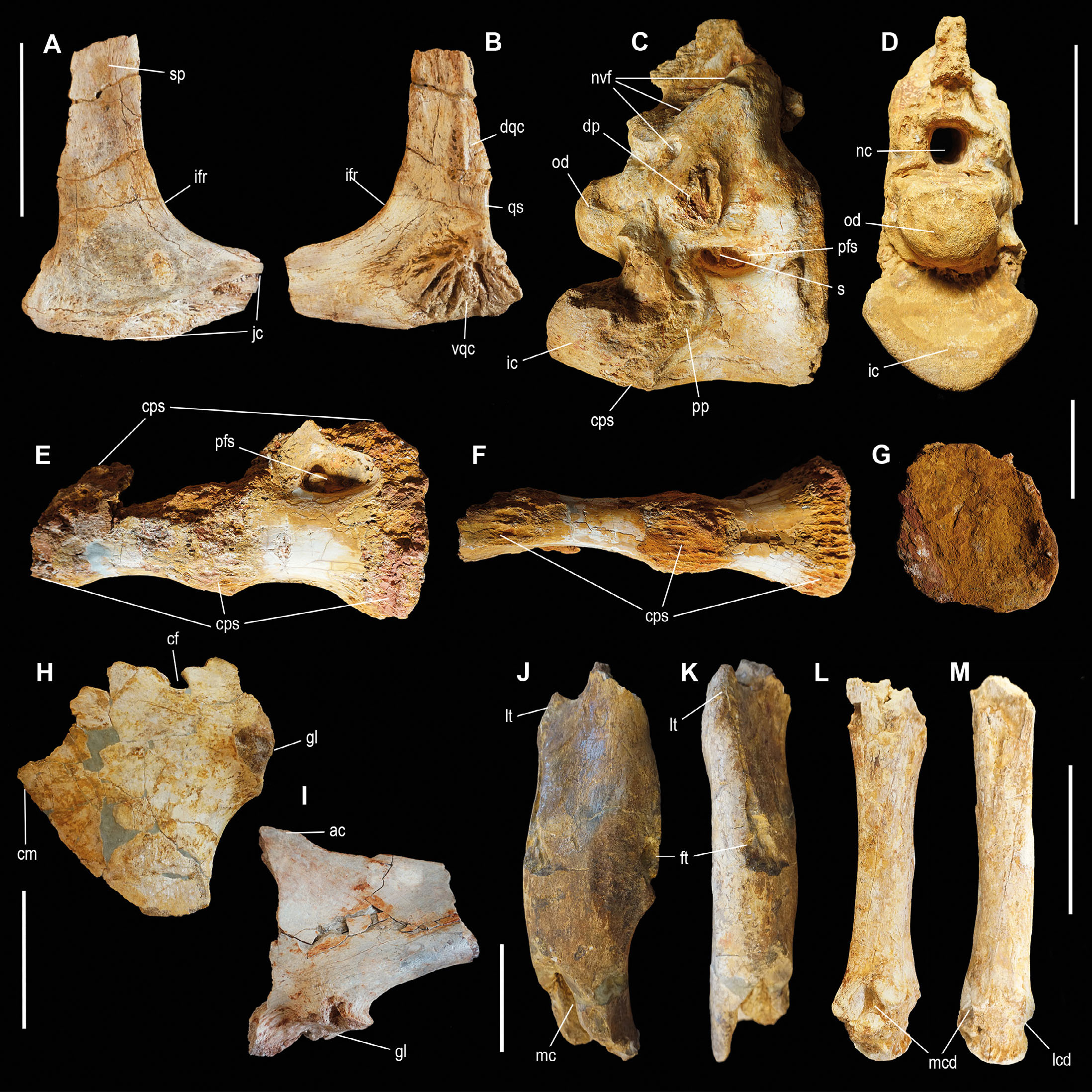
The overall size of the bone is comparable to those of the mid-sized theropods (e.g., Irritator holotype, Schade et al., 2023), yet its ontogenetic status is unknown. It is referred to Averostra incertae sedis.
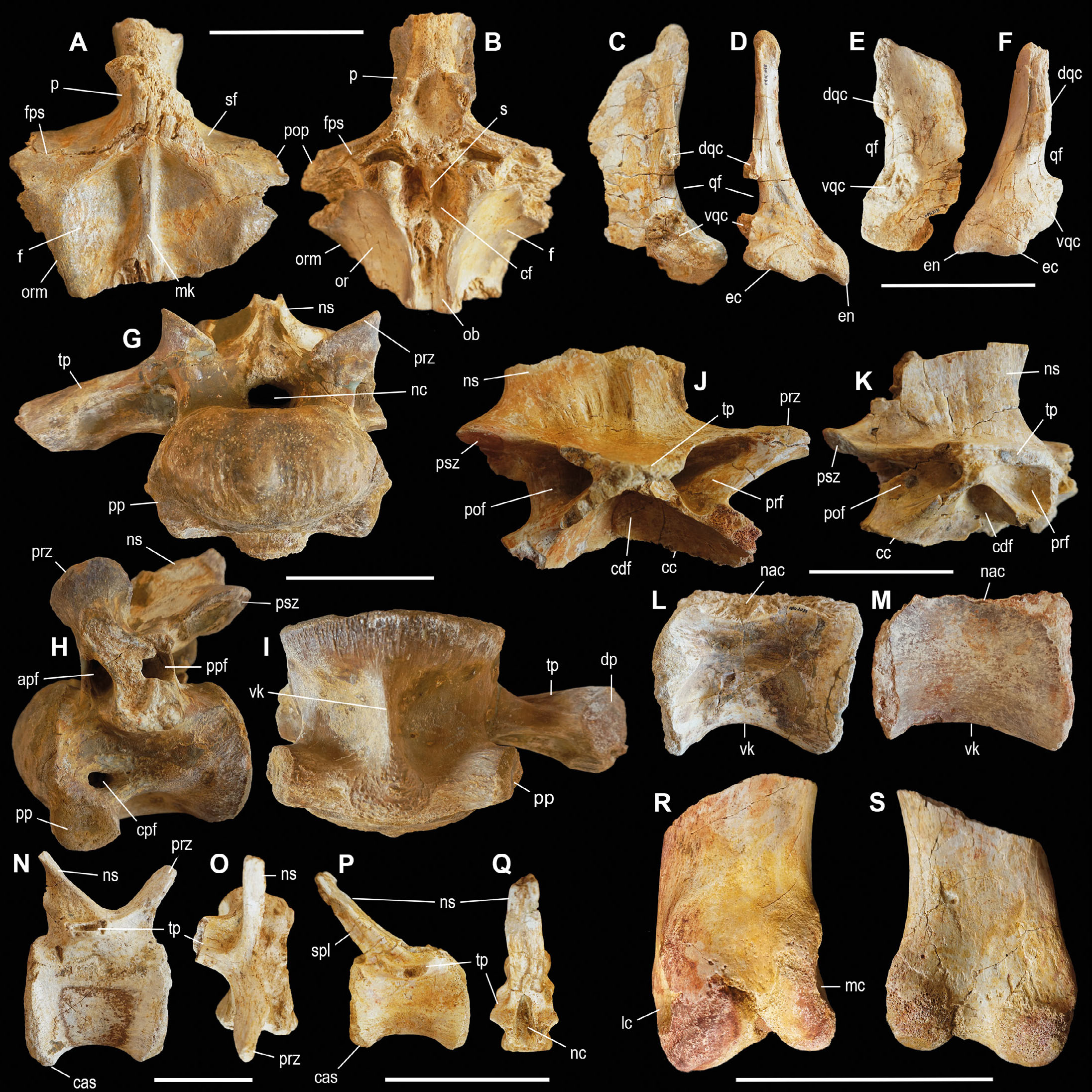
Sacral vertebrae. The specimen OPH2120 is a partial sacrum (Fig. 4E-G). The specimen is composed by two distinct vertebrae, plus a fragment of a third one (the most anterior) all firmly fused together, for a total length of 34.5 cm. Based on comparison with other large-bodied theropods (e.g., Tyrannotitan, Canale et al., 2015), we infer positions 3rd to 5th for the preserved elements of the sacrum. In the eroded areas of the periosteum, the centrum pneumatisation is exposed. It shows a dense pattern of trabeculae and camellae, some of which are encrusted by crystals. The ventral surface is the best preserved part of the vertebrae. The two most preserved vertebrae (S4 and S5) are ventrally concave and both have a narrower anterior facet which in both cases is about 41% the size of the posterior facet, suggesting that the complete sacrum was transversely narrower in the middle region than in the posterior end. In left lateral view, the dorsal half of the lateral surface of the most posterior centrum shows a pneumatic foramen surrounded by a large shallow fossa. In overall features, OPH2120 closely recalls the sacrum of the South American carcharodontosaurid Tyrannotitan chubutensis MPEF-PV1157 (Canale et al., 2015).
The phylogenetic analysis supports two main alternative placements for OPH2120 (marked by “p” in Fig. 3): among early-diverging ceratosaurs in “Abelisauroid clade 1”, and among Carcharodontosauridae. It is provisionally referred to Averostra incertae sedis.
Pectoral girdle elements. The specimens OPH2233 and OPH2234 include two non-associated elements of the pectoral girdle, a left coracoid and the proximal portion of a left scapula (Fig. 4H-I). The coracoid OPH2234 is partially preserved, lacking the scapular margin and an anterodorsal section. The external surface is smooth, medially concave and laterally convex, and bears the glenoid facet and supracoracoid nerve foramen. There is no evidence of a distinct coracoid tuber on the lateral surface. The left scapular fragment, OPH2233, lacks most of the scapular blade, the anterior margin and the tip of the acromion. The preserved element is laterally convex, medially concave and curves along its proximodistal axis. Both specimens show overall similarity with pectoral elements of Baryonyx BMNH R9951 (Charig & Milner, 1997) and Iberospinus ML1190 (Mateus et al., 2011) which might indicate spinosaurid affinities. The absence of the coracoid tuber and the relatively shallow curvature of the posteroventral margin in OPH2234 recalls Bahariasaurus (Stromer, 1934; Sereno et al., 1996). The posteroventral margin of the shaft just distal to the glenoid in OPH2234 differs from the broader arch in Bahariasaurus (Stromer, 1934; Sereno et al., 1996). The material is too fragmentary to be included in the phylogenetic analyses and is provisionally referred to Averostra incertae sedis.
Femur. The specimen OPH2107 consists in the proximal part of the diaphysis of a right femur (Fig. 4J-K). The bone is proximally interrupted at the base of the neck and at the apex of the anterior trochanter. The fourth trochanter is well-preserved. Distally, the specimen is broken off and exposes the medullary cavity. Proximally, the eroded section of the femoral neck is cancellous, and is characterised by a texture similar to those found in the nasal fragment OPH2212, in the cervical vertebra OPH2208, and in the sacral vertebrae OPH2120. The distal portion of the diaphysis, beyond the base of the fourth trochanter, assumes a cylindrical shape that curves caudally.
The material is too fragmentary to be included in the phylogenetic analyses.
Metatarsal II. The specimen OPH2213 is a left metatarsal II (Fig. 4L-M). The fossil is essentially complete, except at the proximal end, where the articular surface is eroded, and distally where the extensor fossa is partially damaged. Proximally, the shaft surface is posterolaterally flattened at the articular contact with metatarsal III. Distally, the shaft bends laterally at an angle of about 7°. The transverse mid-shaft diameter of the bone is about 17% of its proximodistal length. The bone shaft is hollow, housing a distinct medullary cavity. Although in overall shape and proportions, the bone is similar to the second metatarsals of many allosauroids, e.g., Allosaurus (e.g., DNM116 / UUVP6000, Rauhut, 2003; USNM8423, Holtz, 1995; DINO11541, Chure & Loewen, 2020), Mapusaurus (e.g., MCF-PVPH-108.34, Coria & Currie, 2006), Acrocanthosaurus (NCSM14345, Currie & Carpenter, 2000), and Meraxes (Canale et al., 2022), it lacks unambiguous synapomorphies of any theropod clade, preventing a more accurate referral.
The material is too fragmentary for being included in the phylogenetic analyses.
Tetanurae Gauthier, 1986
Axis. This specimen, OPH2025, is a second cervical vertebra of very large size and stout proportions (Fig. 4C-D). In overall morphology, the specimen shares most of its traits with the second cervical of Acrocanthosaurus atokensis SMU746463B-1 described by Harris (1998). The vertebra is laterally compressed and opisthocoelus, firmly fused with the intercentum and the odontoid. The intercentrum is exactly half the length of the vertebral body, extending for a third of the total length of the specimen. This is surmounted dorsally by the odontoid, which appears as a hemispherical projection, slightly compressed vertically, which protrudes anteriorly for the posterior two thirds of the intercentrum. Above the intercentrum, the neural canal is circular in shape, with a diameter of 30 mm. The neural arch is incomplete, lacking the epipophyses and the spinous process. The neural arch is truncated at the base of the postzygapophyses and at the insertion site of the interspinous ligament scars. The two prezygapophyseal-spinodiapophyseal fossae at the base of the prezygapophyses are still preserved. Laterally, just below the eroded base of the diapophysis, an elliptical pneumatic fossa is extended sagittally. Pneumatic foramina open inside these fossae: the one located on the left fossa is divided by a septum (as in OPH2208, Mapusaurus and Acrocanthosaurus atokensis, Harris, 1998; Canale et al., 2015). Two foramina are placed anterodorsally to these fossae: a larger foramen is situated between the diapophyses and the prezygapophyses, and a second one at the base of the prezygapophyseal-spinodiapophyseal fossa. The ventral surface of the centrum bears a longitudinal central ridge, slightly concave in lateral view, narrower at the contact with the intercentrum and wider towards the posterior margin of the vertebral body. Anteroventrally, at the sutural contact with the odontoid, several camellae are exposed on the eroded centrum surface.
Although the specimen closely recalls the axis of the carcharodontosaurids in both size and morphology (supporting such referral following the apomorphy-based approach), the phylogenetic analysis reconstructed two alternative placements for OPH2025 (marked by “o” in Fig. 3) as, respectively, an acrocanthosaurine carcharodontosaurid or a megaraptoran. Accordingly, it is provisionally referred to Tetanurae incertae sedis.
Femur. The specimen OPH2323 is a partial left femur of very large size (Fig. 5F-K). The specimen is missing the proximal end and all of the bone distal to the fourth trochanter. The proximal portion of both neck and anterior trochanter are preserved. The posterior margin of the shaft, lateral to the fourth trochanter, is eroded, revealing the internal texture of the bone. The distal margin shows a dense compact bone along the whole section, which is wider in the lateral and medial sides, reaching a bone depth of 50 mm. Occurring in proximity of the epiphysis, the section in the proximal margin appears porous internally, while an external layer of compact bone surrounds it in correspondence of the lesser trochanter and the femur’s head, where the bone’s section reaches its deepest traits with 70 mm of depth. The suboval medullary cavity occupies about 23-25% of the exposed surface of the bone, a value comparable to many non-theropod dinosaurs, and intermediate between those of most theropods and the very reduced medullary cavity of some spinosaurids (Fabbri et al., 2022). The shaft of the bone distal to the fourth trochanter is elliptical, with an eccentricity (mediolateral diameter vs anteroposterior) of about 1.3, a value higher than those of many large-bodied theropods (e.g., Acrocanthosaurus, Bahariasaurus, spinosaurids: see Carrano, 2006), but smaller than that of some carcharodontosaurids (e.g., Tyrannotitan; Canale et al., 2015). The base of the neck is aligned with the long axis of the shaft cross section, suggesting that the head was medially directed relative to the mediolateral axis of the distal end, as in Bahariasaurus and most tetanurans (e.g., Mapusaurus, Coria & Currie, 2006). The base of the anterior trochanter is robust and forms a distinct projection from the shaft, as in abelisauroids and most tetanurans (e.g., Carrano et al., 2002; Coria & Currie, 2006). The fourth trochanter is placed along the medial margin of the posterior surface of shaft, with its proximal margin immediately distal to the trochanteric region. The fourth trochanter is moderately developed and flange-like with proximal and distal margins gently confluent with the shaft. The perimeter of the shaft distal to the fourth trochanter is 545 mm long, a value comparable to those of the largest carcharodontosaurids and tyrannosaurids (Persons et al., 2020). Using the equation of Campione & Evans (2020), this perimeter suggests the body mass of the specimen at 7.1±1.8 tons, resulting it among the largest known theropods so far described (Persons et al., 2020). This value is the upper limit of the actual body size range inferred for the specimen, because the mid-shaft circumference (the measurement used by Campione & Evans, 2020) is expected to not exceed that taken immediately distal to the trochanteric region.
- Tetanuran material from the Kem Kem Assemblage. Cervical vertebra OPH2208 referred to Carcharodontosaurus saharicus in right lateral (A) and posterior (B) view. Spinosaurid proximal caudal vertebra OPH2123 in left lateral (C), dorsal (D) and ventral (E) view. Tetanuran left femur OPH2323 in anterior (F), medial (G), posterior (H), lateral (I), distal (J) and proximal (K) view. Scale bars = 100 mm. Abbreviations: cas, chevron articular surface; cps, camellate pneumatic structure; ft, fourth trochanter; lt, lesser trochanter; mc, medullary cavity; fs, pneumatic fossa; nac, neural arch contact; prz, prezygapophysis; psz, postzygapophysis; s, septum; tp, transverse process.
The specimen is referred to Theropoda based on the combination of large medullary cavity, prominent anterior trochanter well-distinct from shaft and flange-like fourth trochanter. Although the size of the medullary cavity is proportionally smaller than in most theropods and comparable to sauropods and ornithopods (Fabbri et al., 2022), this feature does not approach some spinosaurids and might be biased by the proximal position of the available cross section, not close to the bone mid-shaft where it is usually sampled. The proximal placement of the fourth trochanter immediately distal to the anterior trochanter, and the symmetric slope of its margins, differentiate this specimen from the femora of both sauropods and large-bodied ornithischians, whose femora have more distally-placed fourth trochanters with different shapes or a lack a distinct muscle insertion (e.g., Norman, 1980; Taylor, 2009). Among theropods, OPH2323 differs from Spinosaurus in the larger size of the medullary cavity and in the shape and lesser development of the fourth trochanter (Ibrahim et al., 2014). The eccentricity of the shaft differentiates it from all carcharodontosaurid femora described in literature (i.e., Acrocanthosaurus, Tyrannotitan), and from Bahariasaurus (Stromer, 1934; Carrano, 2006; Canale et al., 2015). The gently rounded anterior margin of the shaft distal to the anterior trochanter differentiates OPH2323 from the more angular shape in most ceratosaurians, including a large femur from the Kem Kem beds (Chiarenza & Cau, 2016).
It is provisionally considered a Tetanurae incertae sedis.
Carcharodontosauridae Stromer, 1931
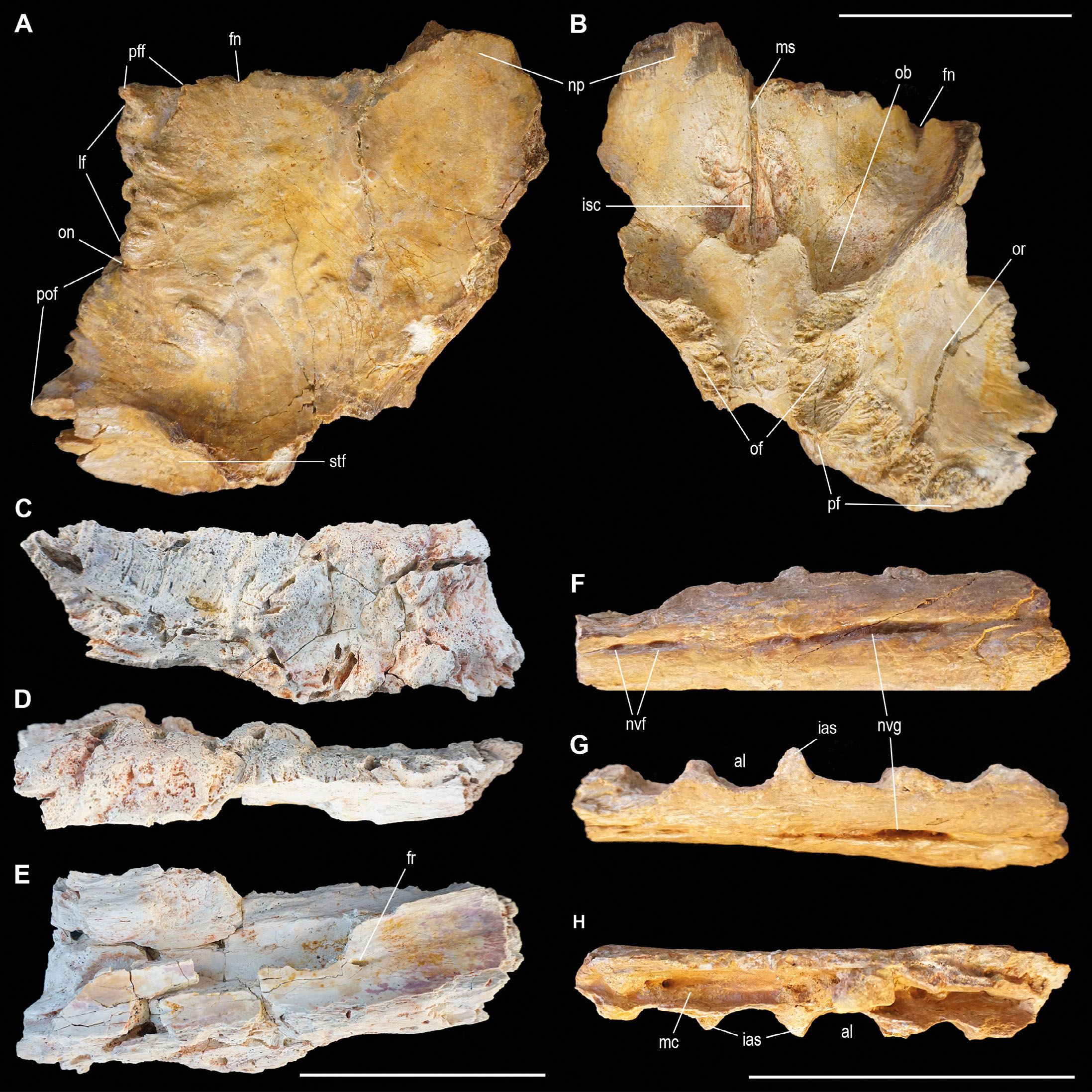
Nasal. The specimen OPH2212 is a thick and compact partial nasal fragment (Fig. 6C-E). The dorsolateral side is the best preserved. Based on comparison with other large-bodied theropod nasals (e.g., Coria & Currie, 2006; Canale et al., 2022), it is likely that the preserved portion corresponds to the posterior half of a right nasal. Along three out of four of the fractured margins, ovoid camellae with a “rice grain” shape are exposed in different concentrations. Dorsolaterally, the fossil is ornamented with a complex hyperplastic texture, shaped by rugosities, furrows, pits and humps as in Mapusaurus (Coria & Currie, 2006, specimen MCF-PVPH-108.1) and Tameryraptor markgrafi, formerly included in the C. saharicus hypodigm (Stromer, 1931; Kellermann et al., 2025, specimen SNSB-BSPG1922X46). The most prominent reliefs are characterised by a porous/spongy surface that includes more or less large pores, while the entire dorsal surface is pierced by microporosity. Also dorsally, within some small cracks, rice grain shaped camellae are present. These are covered by a very thin layer of bone, so to sound empty once subjected to a delicate percussion, especially at the level of the more protruding excrescences. Ventrally, a small foramen is present, similar to those in T. markgrafi (Stromer, 1931; Kellermann et al., 2025). Comparable foramina are found in the nasals of other allosauroids such as Allosaurus fragilis (Snively et al., 2006, specimens UUVP1663 / UMNHVP9146 and UUVP10854 / UMNHVP7784), Mapusaurus (Coria & Currie, 2006), Meraxes (Canale et al., 2022), and Acrocanthosaurus (Eddy & Clarke, 2011, specimen NCSM14345). The ventral surface is smooth, thus even if fragmented, strongly recalling the nasal of Tameryraptor from Egypt (Stromer, 1931). The ruptures expose a large hollow region, which appears elliptical and dorsomedially compressed, extending longitudinally for about 11 cm. Morphological similarities in the section of this fossil are visible in the CT scans of the nasal of Gorgosaurus libratus specimen TMP86.144.1 (Snively et al., 2006). In proximity of the fracture of the ventral plane, the nasal cavity narrows and describes an ellipse. Three foramina are arranged along the closure curve. On the opposite side instead, the cavity reaches its apex of expansion in line with the fracture representing the longitudinal limit of the fossil, where the external edge of the nasal cavity hunches over the lateral wall.
- Carcharodontosaurid material from the Kem Kem Assemblage. Fused frontals OPH2211 referred to cf. Sauroniops sp. in dorsal (A) and ventral (B) view. Right nasal OPH2212 in dorsal (C), lateral (D) and ventral (E) view. Left dentary OPH2026 in lateral (F), dorsal (G) and ventral (H) view. Scale bars = 100 mm. Abbreviations: al, alveolus; fr, foramen; fn, frontal notch; ias, intralveolar septum; isc, contact for the interorbital septum; lf, lacrimal facet; mc, Meckelian canal; ms, medial suture; np, nasal process; nvf, neurovascular foramen; nvg, neurovascular groove; ob, olfactory bulb impression; of, orbitosphenoid facet; on, orbital notch or, orbit; pfc, prefrontal facet; pf, parietal facet; pff, prefrontal facet; pof, postorbital facet; stf, supratemporal fossa.
The material is too fragmentary to be included in the phylogenetic analysis.
Dentary. The specimen OPH2026 is a partially-preserved left dentary (Fig. 6F-H). The distance of the neurovascular sulcus from the alveolar margin, along with the elliptical and anteroposteriorly elongated morphology of the alveoli in dorsal view, suggest that the fragment belongs to the posterior portion of the dentary (see Tyrannotitan MPEF-PV1157, Canale et al., 2015; Giganotosaurus MUCPv-Ch-1, MUCPv-95, Calvo & Coria, 1998; Coria & Salgado, 1995). Laterally, the alveolar margin is straight and the neurovascular sulcus runs parallel to it. The sulcus is uninterrupted in the posterior two-thirds of the fragment, while anteriorly it stops for about 2 cm, resuming with two nutritional foramina in contact with each other.
In ventromedial view, laterally in contact with the alveolar walls, the Meckel’s canal is present along the whole length of the fossil. Such canal is separated from the alveoli by a thin bony wall which thickens in correspondence of the five interalveolar septa. Four preserved alveoli are present, and a first and last incomplete ones are situated at the broken ends. The mesiodistal diameter of the preserved alveoli ranges from 20 to 25 mm. Comparing the alveolar size to those reported in Giganotosaurus MUCPv-95 (25 to 35 mm), and the Moroccan carcharodontosaurid specimens NMC41859 and FSAC-KK02 (Ibrahim et al., 2020a), it can be estimated that OPH2026 belongs to a relatively smaller individual.
The material is too fragmentary to be included in the phylogenetic analyses.
cf. Carcharodontosaurus Stromer, 1931, sensu Kellermann et al., 2025
Cervical vertebra. The specimen OPH2208 is an incompletely preserved cervical vertebra (Fig. 5A-B). The vertebra is sagittally deformed at about 45° towards the left side, and consists only of the vertebral body. The centrum is opisthocoelous, mostly eroded on the left side, yet better preserved in the posterior facet, which appears circular and deeply concave, and in the right lateral surface. The lateral surface bears a distinct pneumatic foramen which is divided into two smaller foramina by a narrow anterior septum. The periosteal surfaces of the parapophysis and along the lipped margin of the posterior facet are bordered by a band of thin transverse ripples. The eroded parts of the centrum show a complex pneumatic structure consisting of trabeculae and camellae that reach and exceed 5 mm in diameter. Despite the missing parts and the taphonomic deformation, the vertebra measures 11 cm along the right side, and 8.8 cm mediolaterally across the preserved portion of the posterior facet.
A similar vertebra, referred to an indeterminate carcharodontosaurid (NMC50792) from the Kem Kem Assemblage was described by Russell (1996) and Evers et al. (2015). This vertebra and OPH2208 are almost identical in the overlapping elements: they share the overall size and proportions, and a similar pneumatisation pattern (including the asymmetric development of the oblique pleurocoelan septum). Accordingly, they have been included in the same OTU. The general morphology, size and pneumatisation pattern of OPH2208 are also very similar to those of the cervical vertebrae of Acrocanthosaurus atokensis (Harris, 1998; Stovall & Langston, 1950), in particular the 5th and 6th vertebrae (SMU746463-54 and SMU746463-66). The proportions of the centrum are also similar to the single known cervical vertebra of Tameryraptor markgrafi, identified as the 4th (Stromer, 1931; Kellermann et al., 2025): the latter differs from OPH2208 in the lateral pneumatic fossa not being divided by an oblique lamina. Overall similarities are also found with the 7th cervical vertebra of Tyrannotitan chubutensis MPEF-PV1157 (Canale et al., 2015). We infer the position of OPH2208 in the cervical series between the 4th and 6th. The specimen NMC50792 shows an unusual feature (McFeeters, pers. com. to AC): a distinct longitudinal ridge running along the roof of the neural canal, and quadrangular, mediolaterally expanded prezygapophseal facets. The former is shared with NMC50403, an isolated posterior dorsal vertebra from the Kem Kem beds (McFeeters, pers. com. to AC; “bone ‘taxon’ C” of Russell, 1996) and with a fragmentary large-bodied theropod from Lybia (Smith et al., 2010; Chiarenza & Cau, 2016).
The coding strings of OPH2208 and NMC50792 were considered part of the same morphotype. The phylogenetic analysis reconstructed this morphotype in the node including Tameryraptor and Carcharodontosaurus. Although the hypodigm of Carcharodontosaurus saharicus sensu Kellermann et al. (2025) currently does not include cervical vertebrae, this morphotype is tentatively referred to the latter species based on the shared geographic and stratigraphic positions.
cf. Sauroniops sp. Cau et al., 2013
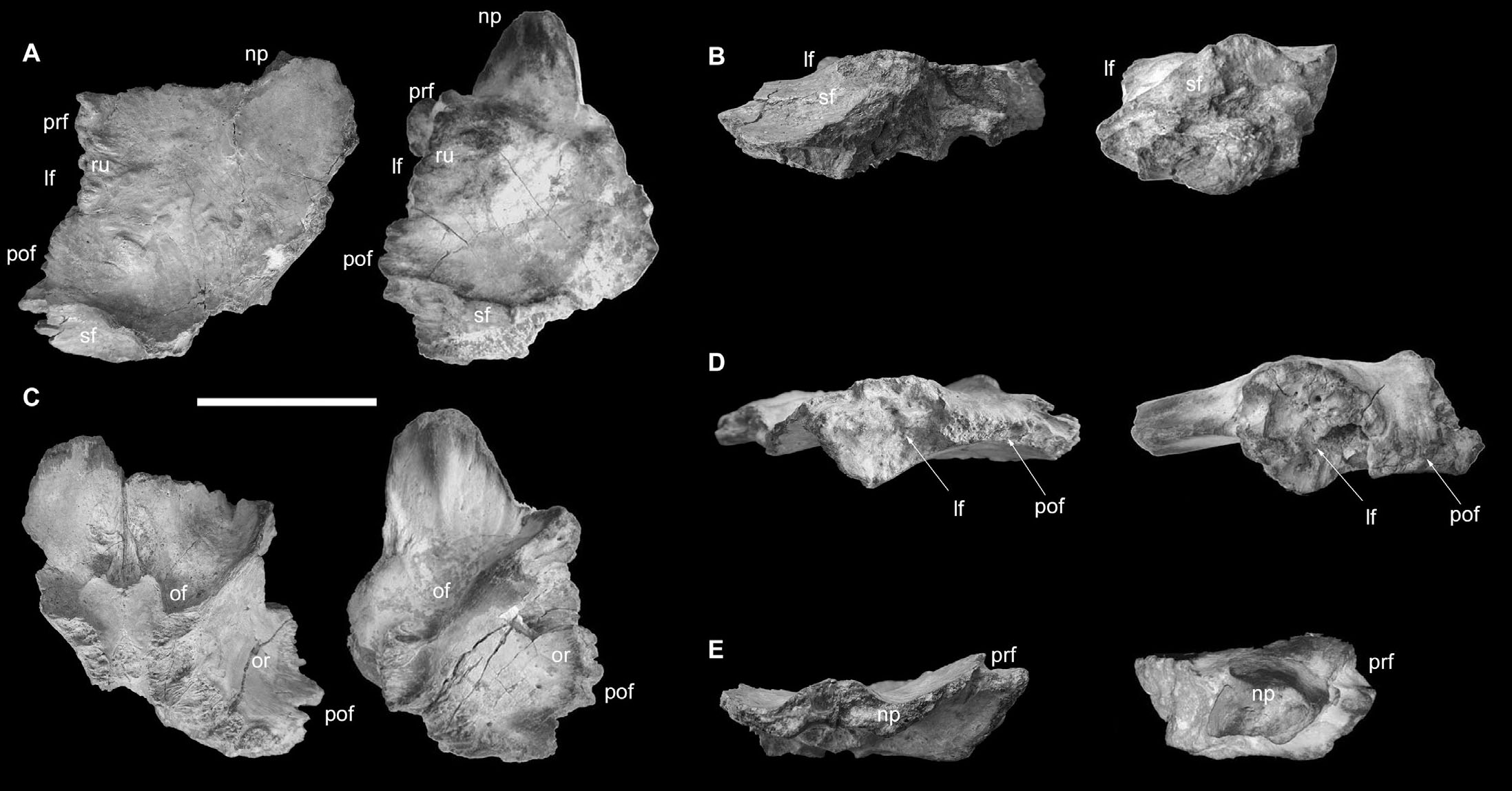
Frontals. The specimen OPH2211 consists in a couple of fused frontal bones of which the left one is the best preserved (Fig. 6A-B). In the right frontal, part of the nasal process is preserved, whereas the prefrontal, lacrimal, postorbital and parietal facets are eroded away. The left frontal, on the other hand, lacks the nasal process, which is truncated at its base, but preserves most of the articular facets with the surrounding cranial bones. The nasal processes seem to be separated medially in correspondence with the fracture of the left one, as in that point, for 1 cm the medial margin of the right nasal process proceeds intact up to its eroded tip. At the lacrimal contact, the fossil reaches its maximum thickness (60 mm), whereas the maximum transversal width of the left frontal is at the posterior half of the postorbital contact (121 mm). The lacrimal margins are laterally raised dorsally, steeply descending medially where they approach the nasal rami, then they raise again in correspondence of the medial suture: such topology generates a small anteromedian ovoid dome. The posterior margin of the lacrimal facet and the anterior margin of the postorbital facet are delimited by a notch, which is especially visible in dorsal view. The prefrontal facet is poorly preserved, and is situated anteromedially to the lacrimal facet, which is separated from the latter by a thin crest from which the two facets are oriented along two distinct inclinations. Ventrally to the shelf-like dorsal prefrontal contact, a second notch is present laterally from the margin of the nasal process. In dorsal view, the nasal rami appear concave from their base. These two concavities rise up medially forming the dome in correspondence with the intranasal suture, and elevate laterally at about 34° from the depression’s lower point to the prefronto-lacrimal contact. The postorbital facet runs transversally in correspondence to the centre of the lacrimal contact: its margins are both dorsally and ventrally concave. The supratemporal fossa is preserved posterior to the postorbital facet. Its anterior margin rises up anteriorly, resulting in a crest which is oriented toward the nasal rami. The dorsal surface of the bone is mostly smooth, but characterised by several rugosities and bumps at the level of the postorbital facet (where the largest rugosities are present), the lacrimal contact, and above the medial portion of the main body (sensu Paterna & Cau, 2022) in correspondence of the medial suture, which is still visible only along its anterior end. Ventrally, instead, such suture is distinct and extends longitudinally from the nasal processes base to the orbitosphenoid contact. The conserved nasal process is convex in ventral view, and shows a slightly visible ventral longitudinal crest. The parietal contact merges with the orbitosphenoid contact generating a crescent suture. The orbital roof is concave, smooth, and pierced by small foramina close to the lateral margin. In ventral view, the fossae for the olfactory bulbs diverge rostrolaterally. Several features of OPH2211 support its referral to Carcharodontosauridae (e.g., Stromer, 1931; Brusatte & Sereno, 2007; Cau et al., 2012, 2013), including the fusion between the two frontals, the presence of both prefrontal and lacrimal facets, the dorsoventrally thickened lacrimal facet, and the posterodorsal orientation of the rostral margin of the temporal fossa. Compared with the two Kem Kem carcharodontosaurids (C. saharicus and S. pachytholus, see Paterna & Cau, 2022), OPH2211 differs from Carcharodontosaurus in the rostromedial placement of the prefrontal facet and in the transversally vaulted dorsal surface (Brusatte & Sereno, 2007), both features shared with MPM2594, holotype of Sauroniops pachytholus (Cau et al., 2013). Yet, despite a comparable longitudinal size (186 mm in MPM2594 vs 170 mm in OPH2211), the specimen differs from the latter in the relatively shallower (dorsoventrally) main body and in the shape of the rostromedial margin of the frontal at the level of the nasal suture, which is dorsally convex in OPH2211 but depressed in Sauroniops (Fig. 8).
The phylogenetic analysis places OPH2211 as sister taxon of Sauroniops. Yet, we do not suggest a species-level attribution since most of the features differentiating OPH2211 from Sauroniops pachytholus holotype (Cau et al., 2012; 2013) could be explained by intraspecific variation, as already shown in other thick-skulled dinosaurs (e.g., Horner & Goodwin, 2009).
Spinosauridae Stromer, 1915
Spinosaurinae (Stromer, 1915)
Skull roof. The specimen OPH2103 is a skull roof composed by both frontals fused with the parietals (Fig. 7A-B). Dorsally, the combined frontals show two symmetrical fossae separated by a medial crest running anteroposteriorly, and posteriorly are delimited by two transversal crests running along the fronto-parietal sutures. The transverse frontoparietal crests join the medial longitudinal crest describing a posteriorly oriented arrow. The two transverse crests gradually merge laterally into the postorbital processes, and posteriorly they delimit the anterior margin of the supratemporal fossae. The parietals are partially preserved and form the thickest part of the frontoparietal complex. Anteriorly, the frontal is truncated at the nasal contacts, with only a small portion of the nasal rami preserved at the anterior end of the medial crest. In the right frontal, the entire prefrontal contact is exposed, whereas in the left element the fronto-prefrontal suture retains a small part of the fused prefrontal. In the frontal, the postorbital and prefrontal facets are separated by a widely arched orbital margin, similarly as in Irritator challengeri (SMNS 58022, Schade et al., 2023), and NHMUKPVR16423 (Lacerda et al., 2024). Lateroventrally, along the orbital margin, a deep groove follows the curve of the orbit and fades at its ends just before reaching the prefrontal and postorbital facets. Ventrally, the orbital roofs are smooth and bound by facets for the postorbital, prefrontal, and orbitosphenoid. The cerebral fossae are well preserved, including the traces of the olfactory bulbs.
- Spinosaurid material from Kem Kem. Skull roof OPH2103 in dorsal (A) and ventral (B) view. Right quadrate OPH2132 in lateral (C) and posterior (D) view. Left quadrate OPH2131 in lateral (E) and posterior (F) view. Cervicodorsal vertebra OPH2011 in frontal (G), left lateral (H) and ventral (I) view. Dorsal vertebra OPH2013 in right lateral view (J). Dorsal vertebra OPH2036 in right lateral view (K). Dorsal vertebra OPH2035 in left lateral view (L). Dorsal vertebra OPH2034 in left lateral view (M). Middle caudal vertebra OPH2214 in right lateral (N) and dorsal (O) view. Distal caudal vertebrae OPH2121 in right lateral (P) and dorsal (Q) view. Pedal phalanx OPH2318 in dorsal (R) and ventral (S) view. Scale bars = 100 mm. Abbreviations: apf, anterior pneumatic foramen; cas, chevron articular surface; cc, centrum contact; cdf, centrodiapophyseal fossa; cf, cerebral fossa; cpf, central pneumatic foramen; dqc, dorsal quadratojugal contact; dp, diapophysis; ec, ectocondyle; en entocondyle; f, frontal; fps, frontoparietal suture; lc, lateral condyle; mc, medial condyle; mk, median keel; nac, neural arch contact; nc, neural canal; ns, neural spine; ob, olfactory bulb; or, orbital roof; orm, orbital rim; pof, postzygapophyseal-centrodiapophyseal fossa; pop, postorbital process; pp, parapophysis; ppf, posterior pneumatic foramen; prf, prezygapophyseal-centrodiapophyseal fossa; prz, prezygapophysis; psz, postzygapophysis; qf, quadrate-quadratojugal foramen; s, septum; sf, supratemporal fossa; spl, spinopostzygapophyseal lamina; tp, transverse process; vk, ventral keel; vqc, ventral quadratojugal contact.
Specimen OPH2103 is similar in size and morphology to the first skull roof morphotype from Morocco defined by Arden et al. (2019), and referred to Spinosauridae (e.g., FSAC‐KK‐3210 and FSAC-KK-7715). The material is coded in “Bone Taxon A” OTU (see Discussion, below) and results nested in Spinosaurinae, in agreement with Arden et al. (2019) and Lacerda et al. (2024).
Quadrates. The two quadrates, specimens OPH2131 and OPH2132, are both well preserved (Fig. 7C-F). The two specimens are of similar size yet differ each other in morphology. Specimen OPH2131 lacks the most dorsal portion, although appearing more massive and lateromedially thickened than the other. The specimen OPH2132 houses a deeper medial fossa running down on an extremely thin wall, and a subtle pterygoid rim. The specimen closely recalls FSAC-KK-18120 (Lakin & Longrich, 2018) and the specimens described by Hendrickx et al. (2016), supporting the referral to Spinosaurinae.
Cervicodorsal vertebra. The specimen OPH2011 is a large theropod cervicodorsal vertebra (Fig. 7G-H, I). It is well preserved with the exception of the left transverse process which is almost completely missing. In overall morphology, it closely recalls the holotype of Sigilmassasaurus brevicollis NMC41857 (Russell, 1996; McFeeters et al., 2013), the specimens BSPG2006I55 (Evers et al., 2015), NMC 41857 (Ibrahim et al., 2020a), and the vertebra referred by Stromer (1934) to “Spinosaurus B”.
The score of the material is redundant with the OTU “Sigilmassasaurus” which is reconstructed among Spinosaurinae in the phylogenetic analysis.
Dorsal vertebrae. The centrum OPH2035 bears both parapophyses and a pleurocentral depression and is interpreted as an anterior dorsal or a cervicodorsal in position (Fig. 7L). In overall morphology, the specimen closely recalls the dorsal vertebrae of the holotype of Spinosaurus aegyptiacus (Stromer, 1915). Both specimens OPH2034 and OPH2035 (Fig. 7L-M) are opisthocoelus and bear a strongly concave caudal articular facet. Both articular ends and the main body of these centra are distincly transversely compressed, hourglass-shaped in dorsal and ventral views. In OPH2034, a prominent medial keel runs along the sagittal axis of the ventral surface. The neural arches (Fig. 7J-K) are similar to those described in the holotype of S. aegyptiacus (Stromer, 1915)
The code strings of this material correspond to various OTSUs of both Spinosaurus type (e.g., char. 1073.1 and 1252.1 of Cau, 2024; cf. plate 1, figure 19 in Stromer, 1915) and neotype (e.g., char. 229.0 and 1252.1 of Cau, 2024; cf. supplementary figure S2E in Ibrahim et al., 2014; see also Sereno et al., 2022).
Proximal caudal vertebra. Specimen OPH2123 is a proximal caudal vertebra (Fig. 5C-E). The vertebral centrum is well preserved except for two erosions, one in the posterior margin, and one in the anterior one on the left side. In lateral view, the anterior and posterior margins of the centrum are delimited by small transverse ridges that follow one another vertically along the entire perimeter of the vertebral body (similarly to OPH2208). In the neural arch, the transverse processes, the spinous process, both prezygapophyses and the right postzygapophysis are missing. The right half of the neural arch is almost completely eroded, except for the base of the postzygapophysis and the medial side of the peduncle. On this side, the cancellous structure of the bone is visible. The left half, on the other hand, still retains the periosteum, with the neurocentral suture completely preserved. The neural canal is intact and unobstructed, appearing circular cranially and elliptical caudally.
The absence of pleurocoentral fossa and pneumatic foramina in the centrum differentiates OPH2123 from both Bahariasaurus and late-diverging carcharodontosaurids (e.g., Stromer, 1931; 1934). The transversal rugosities bordering the intercentral facets recall those in the cervical vertebrae referred to Sigilmassasaurus but unreported in the presacral vertebrae of Spinosaurus (e.g., McFeeters et al., 2013; Evers et al., 2015). Although the camellate-like texture of the bone is unlike those in spinosaurid presacral vertebrae (see above), it is tentatively referred to Spinosaurinae based on overall proportions and the large size (Stromer, 1915; Ibrahim et al., 2020b).
Middle and distal caudal vertebrae. The mid-caudal vertebra OPH2214 includes the vertebral body and the right half of the neural arch, the latter bearing the prezygapophysis, most of the transverse processes and the base of the spinous process (Fig. 7N-O). In posterior view, the neural arch is vertically truncated, lacking the postzygapophyses and the distal part of the neural spine. The distal caudal OPH2121 is well-preserved. Both specimens closely recall the vertebrae of the caudal series of Spinosaurus aegyptiacus FSAC-KK11888 (Ibrahim et al., 2014; Ibrahim et al., 2020b; S. Maganuco, pers. com. to AC, 2022).
The material is an OTSU of the Kem Kem spinosaurine OTU based on FSAC-KK11888.
Pedal phalanx. A large phalanx, OPH2318 (Fig. 7R-S), is interpreted as the second element of the second right toe of a giant-sized theropod based on the combination of dorsoventral shaft compression, flattened ventral surface, asymmetric development in both distal condyles and collateral fossae, presence of a distinct intercondylar sulcus and lack of a distinct extensor fossa (e.g., Brochu, 2003; Ibrahim et al., 2014). The specimen is almost completely preserved, only missing the proximal end. The phalanx is comparable in width and shaft elongation to the homologous element in adult specimens of Tyrannosaurus (e.g., Brochu, 2003), yet it is proportionally shallower along the extensor-flexor axis, differing from the thicker shaft proportions widespread in the pedal phalanges among large-bodied theropods (e.g., tyrannosaurids, e.g., Tyrannosaurus, Brochu, 2003; and carcharodontosaurids; e.g., Meraxes; Canale et al., 2022). In extensor/flexor view, the shaft is moderately constricted relative to the articular ends. The distal articular end is dorsoventrally shallow and bears a wide intercondylar fossa running from the flattened extensor and flexor surfaces. The intercondylar fossa is crossed by a narrow sulcus inclined obliquely. The collateral sulci are elongate proximodistally and occupy most of the corresponding surfaces of the condyles. The medial collateral fossa is wider and better defined than the lateral fossa. The medial condyle is smaller and less extended distally than the lateral condyle. The distal articular surface is wider than deep, with the dorsoventral diameter being 65% of the mediolateral diameter. Using the distal articular facet of this phalanx as a proxy of the proximal facet of the distally-articulating phalanx (the second toe ungual), we suggest that the proximal facet of the second pedal ungual was wider than deep. Among large-bodied theropods, such proportions of the pedal unguals are diagnostic of Spinosaurus (e.g., Maganuco & Dal Sasso, 2018). In the neotype of Spinosaurus, the proximal width of the pedal unguals is consistent along the four toes (see table 1 in Maganuco & Dal Sasso, 2018). The mediolateral width of the toe ungual inferred for the individual represented by OPH2318 is about 145% larger than the values in the toe unguals of the subadult Spinosaurus neotype: this gigantic body size is in agreement with those inferred for the largest spinosaurine specimens (e.g., MSNMV4047, see Maganuco & Dal Sasso, 2018).
The material is too fragmentary to be included in the phylogenetic analysis.
This study also investigated the affinities of other fragmentary theropod specimens from the “mid-Cretaceous” of northern Africa, described in literature but rarely included in numerical analyses.
Smith et al. (2010) described a fragmentary large-bodied theropod from the Aptian-Albian of Lybia and referred it to the ceratosaurian clade Abelisauroidea. The analysis performed here placed it as sister taxon of the penecontemporary Kiyacursor among earliest-diverging abelisauroids.
Mahler (2005) described an isolated maxilla from the Kem Kem units (UCPC10), and referred it to Abelisauridae. Ibrahim et al. (2020a) suggested the referral to Rugops. The analysis performed here placed this specimen in a branch of Abelisauridae including Rugops, albeit closer to another maxilla from South America (Lamanna et al., 2002) and an abelisaurid from India (Dalman & Gislick, 2011).
Salem et al. (2022) described an isolate cervical vertebra from the Bahariya Formation and referred it to Abelisauridae. Our analysis confirms this interpretation and places the specimen in an abelisaurid subclade distinct from the other African abelisaurid OTUs included in the taxon sample.
DISCUSSION
The status of the skull roof morphotypes “Bone Taxon A”, “Morphotype A” and “Morphotype B”
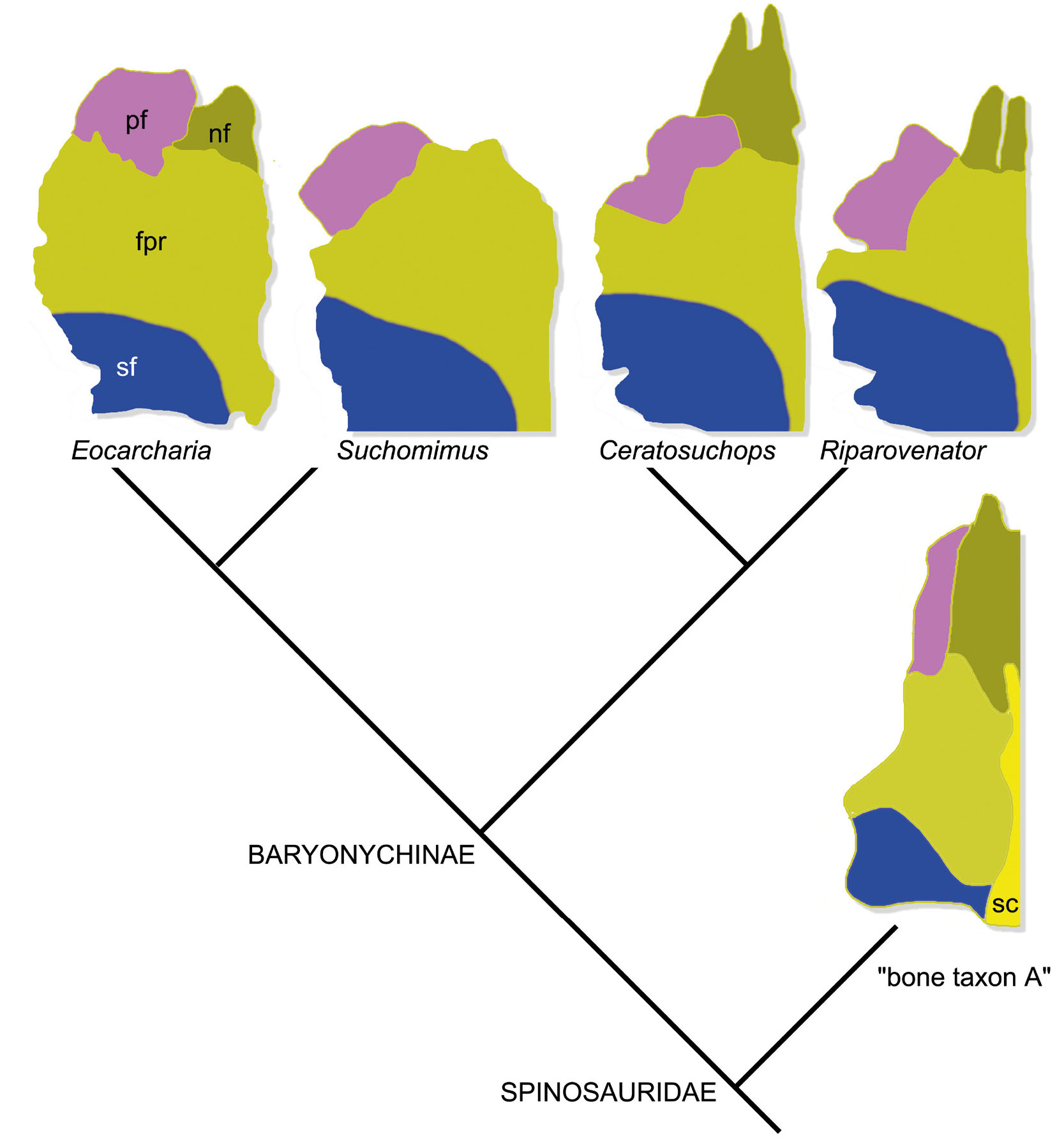
Russell (1996) described two fragmentary skull roof elements from the Kem Kem assemblage and referred them to a theropod “Bone taxon A” of uncertain affinities. Dal Sasso et al. (2005) noted that the ventral surface of “Bone taxon A” matches the posterior end of a partial nasal they referred to cf. Spinosaurus. Arden et al. (2019) described a series of isolated theropod skull roofs from the Kem Kem assemblage and referred them to two spinosaurine morphotypes: they tentatively referred the smaller and more gracile morph (“morphotype A”) to Spinosaurus, and the larger and more robust (“morphotype B”) to Sigilmassasaurus. Lacerda et al. (2024) described a partial skull roof from the Kem Kem assemblage and noted that in both size and morphology it is intermediate between the two morphotypes of Arden et al. (2019), a conclusion which challenges the referral to two distinct taxa by the latter authors. We note that both morphotypes of Arden et al. (2019) and the specimen described by Lacerda et al. (2024) share several features which define “Bone taxon A” of Russell (1996): triangular coossified frontals with narrow endocranial tract delimited by distinct ventral ridges, large participation of the frontal to the orbital margin, vaulted frontal body in lateral view, and fused parietals bearing a sagittal crest. The four morphotypes differ each other mainly in size-related features reported in ontogenetic series of large-bodied theropods, such as length-to-width ratio of the roof, bone thickness, and depth of the articular facets (see Yun et al., 2022, contra Arden et al., 2019), with the specimens described by Russell (1996) being the smallest (about 60% the width of “Morphotype B”; Arden et al., 2019) and, likely, the most immature. We thus consider all these frontoparietal morphologies as semaphoronts of the same morphocline, and refer them to a single taxonomic unit (“Bone taxon A”). The phylogenetic analysis confirms the spinosaurine status of this morphotype (Dal Sasso et al., 2005; Arden et al., 2019).
Chimaerical status of Eocarcharia hypodigm and comments on the inclusiveness of Ceratosuchops and Suchomimus
Recently, Kellermann et al. (2025), following Schade et al. (2023), suggested that part of the material referred to Ceratosuchops (Barker et al., 2021) and Suchomimus (MNNGAD302; Sereno et al., 2022) pertain to carcharodontosaurians, and might be referred to, respectively, Neovenator and Eocarcharia. We disagree with both suggestions. First, note that we agree with Kellermann et al. (2025) and consider the maxilla referred to Eocarcharia as an OTU distinct from the rest of the material (i.e., the skull roof and the holotypic postorbital), but disagree in referring to the latter taxon the specimen illustrated by Sereno et al. (2022). Although similar to Eocarcharia in overall features, the postorbital included in the latter material (see appendix 5 - fig. 2 in Sereno et al., 2022) differs in lacking a suborbital process distinct from the rest of the orbital margin, and in lacking the twisting of the jugal facet onto the lateral surface at the level of the suborbital process (Sereno & Brusatte, 2008). Furthermore, the prefrontal of Eocarcharia is proportionally broader, the prefrontal facet of the frontal is oriented anteriorly and is “V”-shaped in dorsal view (Sereno & Brusatte, 2008), contrasting with the linear and anterolaterally-oriented facet visible in the specimen illustrated by Sereno et al. (2022). In support of the referral of the latter specimen to Suchomimus, the skull roof illustrated by Sereno et al. (2022) shows similarities with both Baryonyx and Irritator in having a trapezoid frontal lacking elongate nasal processes, in bearing a “U”-shaped nasal facet extended posteriorly over the frontal body (Schade et al., 2023), and in showing a mediolaterally broad and ovoid dorsal surface of the prefrontal lacking an anterior ramus (see Charig & Milner, 1997; supplementary material in Barker et al., 2021).
There is no evidence that the Ceratosuchops hypodigm is chimaerical, and we consider very unlikely that both hypodigms of Ceratosuchops and Riparovenator resulted by the accidental co-occurrence of baryonychine premaxillae and basicrania combined with carcharodontosaurian skull roofs (as suggested by Schade et al., 2023). The postorbital of Ceratosuchops shares with that of the spinosaurid Irritator a downturned anterior end of the anterodorsal process and a “U”-shaped-cross section of the ventral process (the latter is a megalosauroid synapomorphy absent in allosauroids; Carrano et al., 2012), but lacks unique allosauroid apomorphies. The features in Ceratosuchops postorbital recalling the carcharodontosaurids are homoplastic among large-skulled theropods (e.g., the rugose and laterally-overhanging supraorbital brow and the ventral ramus bearing a suborbital process are both present also in tyrannosaurids and abelisaurids; Brochu, 2003; Sampson & Witmer, 2007), thus cannot unambiguously support a carcharodontosaurian status. The similarities between Eocarcharia and both Ceratosuchops and Riparovenator, noted by Schade et al. (2022) could thus be explained, alternatively to the latter authors, referring the former genus to the clade containing the latter two taxa (i.e., Spinosauridae). Part of the hypodigm of Eocarcharia shows several spinosaurid-like features which are absent in carcharodontosaurids (Sereno & Brusatte, 2008): 1) the frontal is vaulted in lateral view and is thicker toward the anterior end (see Schade et al., 2024); 2) the anterior end of the frontal is broadly overlapped by the nasal, the latter extended posterior to the level of the anterior margin of the prefrontal (Barker et al., 2021; Lacerda et al., 2024); 3) the prefrontal is dorsally broad, thick and bears a rugose boss (Barker et al., 2021; Lacerda et al., 2024); 4) the prefrontal facet of the frontal is mediolaterally expanded, placed anterolaterally and not reduced in size (Barker et al., 2021; Schade et al., 2024); 5) the lateral margin of the supratemporal fossa of the frontal is notched in dorsal view (Barker et al., 2021; OPH2103); 6) the olfactory tract is very narrow and bound by distinct ridges (Lacerda et al., 2024; OPH2103); 7) the anterolateral ramus of the parietal forms most of the supratemporal fossa excluding the frontal from the anterior margin of the temporal fenestra (Barker et al., 2021; Lacerda et al., 2024; OPH2103); 8) the postorbital facet of the frontal is thickened and houses a distinct sulcus (Barker et al., 2021); 9) the postorbital bears a jugal facet which twists from the posteromedial to the lateral surface along its proximodistal extent (Barker et al., 2021). The first seven features are shared with the spinosaurids but are absent in carcharodontosaurians (e.g., Brusatte & Sereno, 2007; Allen et al., 2019; Schade et al., 2023), the latter two - considered autapomorphies of E. dinops by Sereno & Brusatte (2008) - are uniquely shared with Ceratosuchops among known theropods (Barker et al., 2021). Differing from all allosauroids retaining the prefrontal (e.g., Allosaurus, Madsen, 1976), Eocarcharia shares with Baryonyx the apomorphic absence of the anteroventral ramus, which in theropods usually participates in the orbital margin (Lacerda et al., 2024), and recalls all baryonychines in having the long axis of the prefrontal oriented obliquely and anteromedially relative to the frontal long axis, instead that parasagitally (cf. Lacerda et al., 2024, with Barker et al., 2021). Given the numerous derived features shared with Baryonyx, Ceratosuchops, Irritator and Riparovenator, the most parsimonious interpretation is to refer the postorbital and the skull roof material of Eocarcharia to a spinosaurid (Fig. 8), distinct from the rest of the hypodigm (i.e., an isolated maxilla, Sereno & Brusatte, 2008). The result of our phylogenetic analysis confirms this interpretation, placing the Eocarcharia COTSU (i.e., based on the skull roof elements and postorbital) among Baryonychinae. Although the name “Eocarcharia dinops” explicitly refers to the carcharodontosaurid maxilla (Sereno & Brusatte, 2008), the holotype (and, thus, the name-bearer specimen) of Eocarcharia dinops is the postorbital included in the OTU which clusters among the ceratosuchopsini baryonychines in our phylogenetic analysis. Accordingly, Eocarcharia dinops should be referred to Spinosauridae and not to Carcharodontosauridae. The most intriguing detail of this novel relationships is the direct sister taxon relationships between Eocarcharia dinops (sensu stricto, see above) and Suchomimus (sensu lato, see Sereno et al., 1998; Barker et al., 2021), both from the Elrhaz Formation of Niger (Sereno & Brusatte, 2008). Pending a revision of the Suchomimus hypodigm, the two OTUs differ in 14 character states, and should be considered distinct taxa. Such result further supports the hypothesis that the co-occurrence of two (or more) closely-related species is a recurrent pattern in spinosaurid evolution (Evers et al., 2015; Barker et al., 2021) and this should not preclude the coexistence of multiple Spinosauridae subtaxa in the ‘Kem Kem beds’ associations (contra Ibrahim et al. 2020; Smith et al. 2020). Sereno et al. (2022) noted that the features differentiating Riparovenator from Ceratosuchops co-occurr in the Suchomimus hypodigm (and, in some cases, even in the same specimen), and suggested to refer the two Isle of Wight baryonychines to a single taxon (Ceratosuchops) (contra Barker et al., 2021). Assuming the distinction of Eocarcharia from Suchomimus among baryonychines, the Suchomimus hypodigm as currently defined (“Suchomimus sensu lato”) could also include yet-unrecognised Eocarcharia material, and thus could incorporate a diversity above the species level. A detailed comparative analysis between the whole Suchomimus (sensu lato) hypodigm and Eocarcharia (sensu stricto) would help elucidating the systematic meaning of the features differentiating Riparovenator and Ceratosuchops.
- Comparison between OPH2211 (left) and Sauroniops pachytholus holotype MPM2594 (right), in dorsal view (A), posterior view (B), ventral view (C), left lateral view (D), anterior view (E). Scale bar = 100 mm. Abbreviations: lf: lacrimal facet, np: nasal process; of: olfactory bulb fossa; or: orbital roof; pof: postorbital facet, prf: prefrontal facet; ru: rugosities; sf: supratemporal fossa.
- Schematic diagram of the articulated prefrontal and frontal bones of selected spinosaurids in dorsal view. Abbreviations: fpr, frontoparietal subcutaneous roof; nf, nasal facet; pf, prefrontal; sc, sagittal crest; sf, supratemporal fossa. Modified from Sereno & Brusatte (2008); Barker et al. (2021); Sereno et al. (2022); Lacerda et al. (2024).
Enforcing our analysis to reconstruct both Eocarcharia OTUs in Allosauroidea requires eight steps more than the shortest unenforced topologies. The isolated maxilla originally referred to E. dinops by Sereno & Brusatte (2008) lacks spinosaurid features, is consistently reconstructed by our analyses in the carcharodontosaurid clade, and should be referred to a new taxon. The postcranial material originally referred to Kryptops palaios by Sereno & Brusatte (2008) is also reconstructed in Allosauroidea, although outside Carcharodontosauridae and among Metriacanthosauridae. This result suggests an Early Cretaceous radiation of the metriacanthosaurids ranging from Europe (Erectopus superbus), Southeastern Asia (Siamotyrannus) and Africa (“Kryptops postcranium”). Alternatively, future discoveries may demonstrate that the two Elrhaz Formation allosauroid OTUs belong to the same taxon and that their distinction was biased by the lack of overlapping material. Based on the available evidence, the scenario enforcing both Eocarcharia OTUs and the postcranium referred to Kryptops to belong to a single species is six steps less parsimonious than the shortest unenforced topologies. The resolution of the taxonomy of the Elrhaz Formation allosauroid material is beyond the scope of this study.
The status of Bahariasaurus and other gracile-limbed abelisauroids
Stromer (1934) described a large collection of mid- to large-bodied theropod remains from the Bahariya Formation of Egypt and erected Bahariasaurus ingens from a fragmentary individual including a few dorsal vertebrae, a partial sacral series and incomplete pelvic elements. Several specimens of various size from the same unit were referred by Stromer (1934) to Bahariasaurus. Since the type and all the material referred to Bahariasaurus was destroyed during WWII and is now lost, the taxon has received very little attention (e.g., Rauhut, 1995), and has been included in numerical analyses only recently. Sereno et al. (1996) moved part of the Bahariasaurus hypodigm (sensu Stromer, 1934) to Deltadromeus. The latter genus has been alternatively considered a coelurosaurian (Sereno et al., 1996; Rauhut, 2003; Cau, 2018; Motta et al., 2016), a ceratosaurian (Sereno et al., 2004; Carrano & Sampson, 2008; Cau, 2024), or an allosauroid (Apesteguia et al., 2016). Carrano & Sampson (2008) questioned the referral of part of the Bahariasaurus hypodigm to Deltadromeus, and noted ceratosaurian-like features in the holotype sacrum. Chiarenza & Cau (2016) and Motta et al. (2016) discussed possible affinities of the “bahariasaurids” (meant to include Deltadromeus), suggesting, alternatively, an abelisauroid or tyrannosauroid status for that group. Cau (2024) included an OTU based on Bahariasaurus but restricted to the type material in a large-scale phylogenetic analysis, where it clustered with Deltadromeus and a few other taxa among an early-diverging lineage of Ceratosauria.
Here, we suggest a solution to the controversial status of Bahariasaurus and Deltadromeus. As described in the Material and Method section (above), an isolate pelvic bone from the Bahariya Formation (i.e., SNSB-BSPG1912VIII82; Stromer, 1934), originally interpreted by Stromer as the pubis of a theropod of uncertain affinities, much closely recalls the theropod ischia and shows the peculiar morphology of the proximal end of the Bahariasaurus holotypic ischium (the latter lacks the distal end; Stromer, 1934). At the same time, the distal end of SNSB-BSPG1912VIII82 is identical to the preserved distal end of Deltadromeus agilis ischium (a bone originally considered the pubis, but later reinterpreted as the ischial foot: the true pubic foot of D. agilis has been found articulated with the gastralia, see Carrano & Sampson, 2008, and Apesteguia et al., 2016, contra Sereno et al., 1996; Ibrahim et al., 2020a). The co-occurrence of autapomorphies of both B. ingens and D. agilis in SNSB-BSPG1912VIII82 supports their referral to the same taxonomic unit (Fig. 1). We suggest that most of the differences between Deltadromeus holotype and part of the Bahariasaurus hypodigm are size- or ontogeny-related (the former is based on an immature individual, Ibrahim et al., 2020a) and are compatible with those observed in other large theropods (e.g., Russell, 1970; Molnar, 1990) with an expected large-scale distribution (Farlow & Pianka, 2002). This interpretation explains the broad overlap between the Deltadromeus holotype and the Bahariasaurus hypodigm suggested by Sereno et al. (1996) and Ibrahim et al. (2020a), and provides the basis for further referral to Bahariasaurus of isolated theropod bones from the Bahariya Formation (see Material and Methods, above).
Our phylogenetic analysis supports the referral of Bahariasaurus to Abelisauroidea, in agreement with the placement of Deltadromeus by Sereno et al. (2004), Carrano & Sampson (2008), Rauhut & Carrano (2016) and Cau (2024). In particular, the analysis supports the close relationship between Bahariasaurus and the yet unnamed “Angeac-Charente taxon” from a rich bonebed from the Jurassic-Cretaceous boundary of France and referred to Ornithomimosauria by Allain et al. (2014; 2022). Most of the ornithomimosaur synapomorphies reported in the “Angeac-Charente taxon” by Allain et al. (2014, 2022) are shared by Bahariasaurus or other gracile-limbed abelisauroids and thus could equally support our hypothesis (e.g., toothless dentary, elongate presacral vertebrae, elongate distal caudal vertebrae with broad and low centra bearing hypertrophied prezygapophyses, straight and gracile humerus, separation of the fibular crest from the tibial proximal end, flattened astragalar facet of tibia, large proximomedial fibular fossa bound by a posterior lamella, wide ascending process of astragalus, flattened pedal unguals bearing a fossa instead of the flexor tubercle; Xu et al., 2009; Rauhut & Carrano, 2016; Cerroni et al., 2019; de Souza et al., 2021). Allain et al. (2022) listed further features of the French theropod dismissing an abelisauroids status, and questioned the abelisauroid affinities of Deltadromeus and Limusaurus. The referral of Deltadromeus to Ornithomimosauria was originally suggested by Rauhut (2003) but so far has never been replicated by other phylogenetic analyses. Yet, we remark that the “Angeac-Charente taxon” as described by Allain et al. (2022) shows abelisauroid synapomorphies absent in the ornithomimosaurs, including a hypertrophied external mandibular fenestra, two pleurocoels in the cervical centra, a scapula particularly short and broad, and the fourth metatarsal much gracile than the third (Carrano & Sampson, 2008; Xu et al., 2009; Rauhut & Carrano, 2016). The rich character sample in our phylogenetic analysis includes all the features discussed by Allain et al. (2022): we tested that alternative placement for Bahariasaurus and “Angeac-Charente taxon” enforcing the latter two in Ornithomimosauria. The shortest topologies reconstructed under such constraint are 11 steps longer than the shortest trees found by the unconstrained analysis, a scenario much less parsimonious than assuming an abelisauroid placement for the two theropods. Pending a detailed description of the “Angeac-Charente taxon” we conclude that the latter and Bahariasaurus are gracile-limbed abelisauroids convergent with some coelurosaurs.
Implications for the “Stromer’s Riddle”
Our analysis confirms the absence of coelurosaurs in the theropod communities from the “mid-Cretaceous” of northern Africa and the dominance of the “triumvirate” formed by Abelisauroidea, Spinosauridae and Carcharodontosauridae in these assemblages (Ibrahim et al., 2020a). The diversity among the sampled northern African theropod assemblages is controversial, and despite the shared presence of the “triuvirate of clades”, it cannot be reduced to a unique and homogeneous community spanning the whole continent. The analysis suggests that the Elhraz Formation from Niger includes at least a gracile-limbed abelisauroid, an abelisaurid, possibly two allosauroids (a metriacanthosaurid and a carcharodontosaurid), and a subclade of baryonychine spinosaurids (including two distinct species). The less studied Eckhar Formation from Niger includes one abelisaurid, one spinosaurid (not included in the analysis, see Sereno et al., 2022), and at least one carcharodontosaurid. The Bahariya Formation includes one early-diverging abelisauroid, one abelisaurid, a carcharodontosaurid and at least one spinosaurine. The Kem Kem Assemblage includes at least one gracile-limbed abelisauroid, an abelisaurid, at least one spinosaurine and two carcharodontosaurids. Using the phylogenetic affinities of the sampled taxa as a proxy of faunal similarity among the formations (Fig. 3), the Eckhar Formation and the Kem Kem Assemblage resulted the most similar, and the Elhraz Formation the most different. Although diachrony likely explains the distinction between the Elhraz Formation and other units (Sereno & Brusatte, 2008), more controversial is explaining the difference between the Bahariya Formation and the other Cenomanian-age units, since the former has been considered coeval and sharing the same genera found in the latter two units (e.g., Brusatte & Sereno, 2007; Ibrahim et al., 2020a).
The abundance of large- to giant-bodied theropods in these units, compared to the relative rarity of non-theropod dinosaurs, is usually termed the “Stromer’s Riddle”, and has been considered alternatively the result of peculiar ecological conditions, a preservation artifact, the effect of selective collection biased by commercial exploitation of the fossiliferous units, or a combination of these factors (e.g., Stromer, 1915, 1934; Sereno et al., 1996; Russell, 1996; Sereno & Brusatte, 2008; Dyke, 2010; Chiarenza & Cau, 2016; Ibrahim et al., 2020a). Recent field analyses have dismissed any significant impact of collection bias in altering the diversity of the Kem Kem faunal assemblage, and support a trophic web based on the rich aquatic productivity in a large deltaic system (Läng et al., 2013). Sedimentological analysis of the teeth-bearing units sampled from the Aptian-Albian of southern Tunisia suggests environmental segregation of the theropod taxa, with abelisaurids and carcharodontosaurids more common in inland, fluvial deposits, and spinosaurids more frequent in estuarine and coastal deposits (Fanti et al., 2014). This scenario is further supported by the morphological and isotopic differences between the teeth of the spinosaurids and those of the other theropods from the Kem Kem beds (Amiot et al., 2010; Ibrahim et al., 2020a). The two above-mentioned cases highlight the importance of an integrated palaeoecological approach in addressing the challenges posed by the peculiar stratigraphic and depositional settings of the northern African theropod record. Chiarenza (2024) remarked that separating different facies into distinct chronological horizons can clarify the palaeoecological paradoxes of the “Stromer’s Riddle”. Yet, the shared occurrence of the three main “triumvirate” groups in all sampled northern African units, despite their different species-level compositions, suggests that the ecological partition among the theropod taxa was not merely a zonation issue and supports a hierarchical macroecological structure regulated by clade-specific evolutionary patterns. Several factors, progressively nested, likely shaped the history of these communities. One factor is the ecological specialisation of each lineage, expressed by their trophic and locomotory autapomorphies. This first level of ecological regulation might explain the vicariance between baryonychines and spinosaurines (Sereno & Brusatte, 2008; Ibrahim et al., 2020a), the abundance of spinosaurids compared to other taxa (e.g., Fanti et al., 2014), but is not sufficient for explaining the co-occurrence of abelisaurids and carcharodontosaurids in all units, clades which apparently shared similar trophic specialisations (e.g., deep oreinirostral skulls with ziphodont dentition, Carrano & Sampson, 2008; Carrano et al., 2012).
The phylogenetic framework resulted here helps explaining the ecological role of the most enigmatic among the giant theropods found in these communities, Bahariasaurus. This genus is nested among a clade (“Abelisauroid clade 1”) which is formed by taxa inferred to have herbivorous or omnivorous ecologies (e.g., Limusaurus, Wang et al., 2017; Berthasaura, de Souza et al., 2021). Assuming a similar trophic regime in Bahariasaurus, this taxon may had played the ecological role occupied elsewhere by other non-theropod clades rarely represented in the northern African units, e.g., the ornithischians, and should be removed from the predatory guilds of these communities (e.g., Holtz, 2021; Cau, 2024). Unfortunately, the currently known specimens of Bahariasaurus from the Kem Kem units are immature (Ibrahim et al., 2020a), and this negatively impacts the identification of species-level autapomorphies eventually differentiating the Moroccan population from the Egyptian one (see Cau, 2024).
The carcharodontosaurian fossil record clarifies well the hierarchically-connected issues expressed by these northern African units. One early-diverging carcharodontosaurian, represented by the maxilla originally referred to Eocarcharia by Sereno & Brusatte (2008), is known in the likely older Eckhar Formation. This early-diverging carcharodontosaurian lineage is not represented in the other northern African units, which instead include three later-diverging branches, leading respectively to Sauroniops, to “Carcharodontosaurus” iguidensis, and to the subclade formed by Carcharodontosaurus (sensu stricto, Kellermann et al., 2025) and Tameryraptor: in this case, phylogenetic and stratigraphic structures agree in explaining the faunal differentiation between the Eckhar Formation and the other units. More complex are the relationships between the carcharodontosaurids from the Bahariya, Elhraz and Kem Kem units. The T. markgrafi - C. saharicus sister-grouping suggests allopatric separation between the Bahariya and Kem Kem faunas. Under such a scenario, the apparently primitive morphology of Tameryraptor interpreted by Kellermann et al. (2025) is instead an autapomorphic condition of the latter taxon due to allopatry. At least three carcharodontosaurid frontal morphologies are documented in the Kem Kem units: Carcharodontosaurus saharicus, Sauroniops pachytholus and cf. Sauroniops (Paterna & Cau, 2022; this study). We suggest that cf. Sauroniops could be stratigraphically-distinct from the other two carcharodontosaurids, based on the rough conglomeratic lithology of the sediment encrusting OPH2211, featuring small spherical concretions known as “kerkoubs” (AP, pers. obs.), typical of the Gara Sbaa Formation (Cailleux & Soleilhavoup, 1976; Sereno et al., 2020a), which can also be found on the surface in the Taouz locality (AP, pers. obs.). This differs from the sediment encrusting the holotype of Sauroniops pachytholus which is a reddish fine-grained sandstone (AC, pers. obs.), more characteristic of the higher parts of the Kem Kem successions (Ibrahim et al., 2020a). Although lithology alone is not a sufficient criterion for assessing a proper zonation, OPH2211 was likely collected from a conglomerate bed from the base of the Gara Sbaa Formation (sensu Ibrahim et al., 2020a) at Taouz locality, and thus could be stratigraphically distinct from Carcharodontosaurus saharicus neotype from the Douira Formation (sensu Ibrahim et al., 2020a) found at Gara Sbaa locality. Following this interpretation, the cf. Sauroniops - S. pachytholus sister group might represent an anagenetic sequence driven by the evolution of a more complex cranial ornamentation. Assuming the two Kem Kem carcharodontosaurids found in sandstone facies as coeval, the peculiar skull roof ornamentation of Sauroniops pachytholus compared to Carcharodontosaurus saharicus might represent a display feature evolved for species recognition among sympatric taxa (Cau et al., 2013; Paterna & Cau, 2022).
CONCLUSION
The revision of the theropod material from the Aptian-Cenomanian of northern Africa and the application of methodological splitting in an updated large-scale phylogenetic analysis of Theropoda support the synonymy between the Moroccan Deltadromeus and the Egyptian Bahariasaurus, and suggest a chimaerical status for the purported carcharodontosaurid Eocarcharia. Our analysis also supports the exclusion of Carcharodontosaurus iguidensis from the latter genus. Yet, the analysis cannot support neither rejects a genus-level distinction of the Egyptian carcharodontosaurid specimen described by Ernst Stromer and recently renamed Tameryraptor markgrafi, neither that of Sigilmassasaurus from Spinosaurus. This result does not necessarily dismiss a species-level distinction between the taxa here reconstructed as sister taxa and showing phylogenetically-significant differences (e.g., Tameryraptor vs Carcharodontosaurus, or the Bahariya Formation Spinosaurus vs the Kem Kem Assemblage Spinosaurus). The validity of Sauroniops and its distinction from the other carcharodontosaurids has been further supported by a new specimen referred to that genus. Yet, the redescription of Carcharodontosaurus saharicus neotype skull roof, so far published only partially, would likely help in resolving the controversy on the number and relationships of the giant allosauroids in the Kem Kem Assemblage.
The theropod diversity in the “mid-Cretaceous” of northern Africa cannot be resolved following “splitting” or “lumping” aprioristic approaches: although we confirm that these communities are formed mainly by the non-coelurosaurian “triumvirate”, the number of species of each “triumvir group” sampled in these units was likely underestimated by the rigid application of an extremely conservative lumping taxonomy. This taxonomic issue is exacerbated by the limited overlap between the material referred to most taxa, and by the poor knowledge of the ontogenetic diversity in non-coelurosaurian theropods. Even more significant than the taxonomic issue, that approach introduced chimaerical OTUs in the phylogenetic analyses, like Eocarcharia as originally conceived, which unpredictably inflate the already high homoplasy of the theropod sample.
Direct overlap of diagnostic elements is the only valid criterion for lumping non-associated material. The mere close phyletic relatedness (e.g., the referral of isolated bones to the same “triumvir clade”) is a criterion not sufficiently solid for preventing potential chimaeras. In the absence of overlap in the sampled specimens, hypodigms based on “methodological splitting” should be preferred as null hypotheses for OTU definition because they prevent topological artifacts biased by the unrecognised inclusion of chimaeras in the taxon sample.
From an ecological perspective, the “Stromer’s Riddle” is likely more apparent than real: trophic and environmental segregation operated between the main theropod clades, reducing direct competition. The generalist and semi-aquatic spinosaurids and the probable omnivorous (if not vegetarian) bahariasaurs likely were not in direct competition with the other giant theropods bearing ziphodont dentitions better adapted to the hypercarnivorous diet. Among-theropod predation probably played a significant role in shaping the trophic web of these communities. The “mid-Cretaceous” northern African dinosaur communities were more disparate yet less aberrant than often assumed.