

INTRODUCTION
This year marks the centenary of the first description of one of the most iconic carnivorous dinosaurs, Velociraptor, made infamous as one of the main “antagonists” in the popular film series “Jurassic Park” and its “Jurassic World” sequels. The first Velociraptor fossil (AMNH 6515) was discovered in 1923 during the second of the Central Asiatic Expeditions of the American Museum of Natural History (AMNH) into the Gobi Desert, led by Roy Chapman Andrews, the legendary American palaeontologist, and explorer that may have inspired the popular movie character Indiana Jones (Romano & Novacek, 2023). This remarkable discovery was made on August 11, 1923, in the Upper Cretaceous deposits of the Djadokhta Formation at the Flaming Cliffs locality (Bayn Dzak, Mongolia). The specimen was collected by Peter Kaisen, a veteran collector and preparator at the AMNH (Granger, 1936) and consisted of a complete –albeit laterally crushed– skull and a manual ungual articulated with the penultimate phalanx from digit III (Fig. 1).
- Skull of AMNH 6515 in right lateral view (a), left lateral side (b) and original drawings by Osborn (1924) (d). Manus claw articulated with the penultimate phalanx from digit III of AMNH 6515 (c).
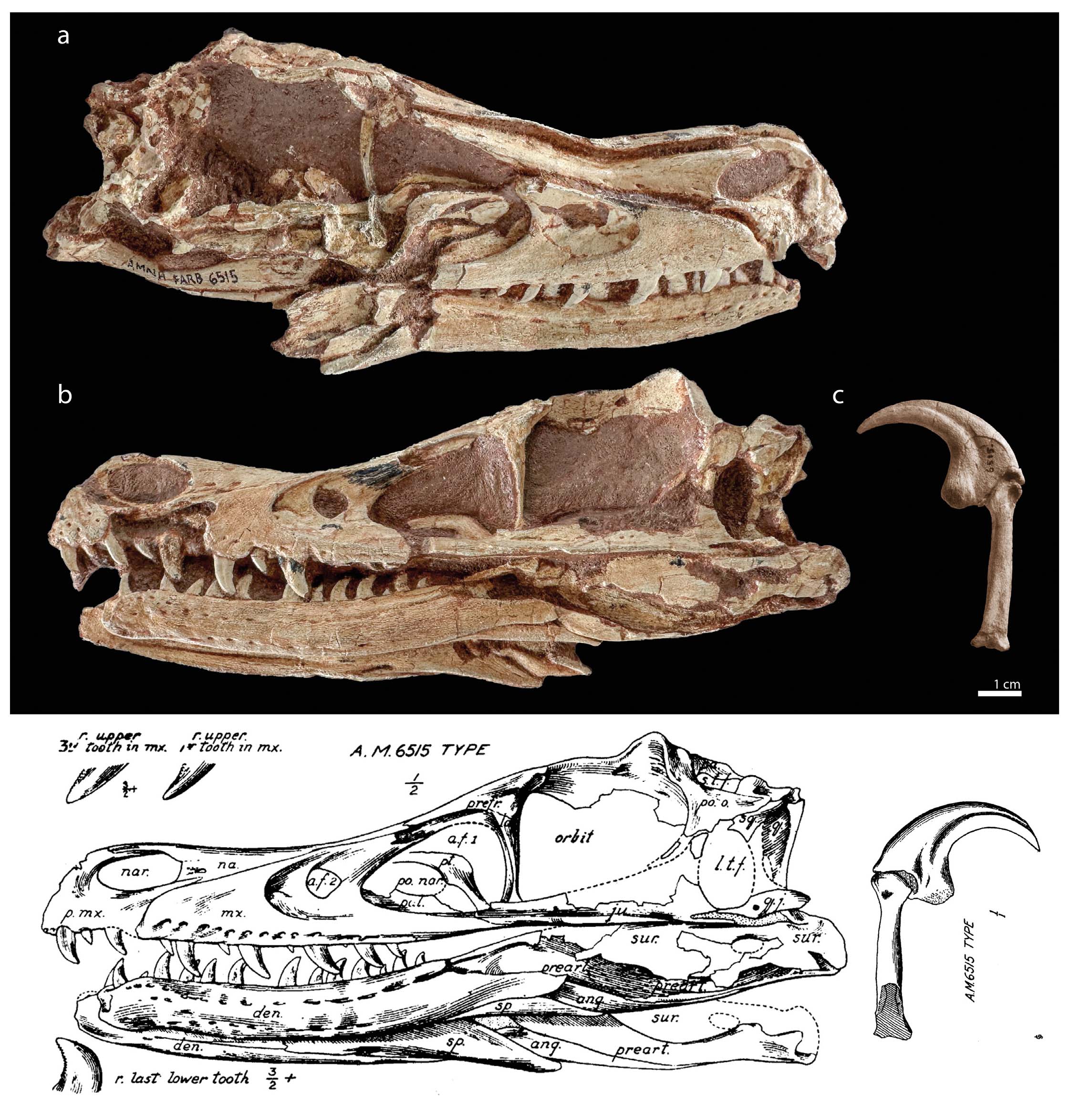
The scientific name of this new dinosaur, Velociraptor mongoliensis, was bestowed a year later by the American palaeontologist and then-President of the AMNH, Henry Fairfield Osborn, in a seminal paper naming three new theropod species (Osborn, 1924). He combined the Latin words “velox” (meaning ‘fast’) and “raptor” (meaning ‘predaceous’ or ‘plunderer’) (Osborn, 1924). The specific epithet “mongoliensis” refers to its source locality in Mongolia. To date, only one other species of Velociraptor is considered valid: V. osmolskae, described in 2008 based on three cranial bones collected at Bayan Mandahu, roughly 300 km southeast of Bayn Dzak (Godefroit et al., 2008), although ongoing statistical research has proposed greater taxonomic diversity among the specimens currently referred to V. mongoliensis (Powers et al., 2020; Ruebenstahl et al., 2023).
Velociraptor and other dromaeosaurids share with other paravians the uses of pedal digits III and IV in locomotion (Li et al., 2007) as suggested by the anatomy of articulated specimens (Norell & Makovicky, 1997) and confirmed by footprints (Li et al., 2007); differently the inner major pedal digit (digit II) is elevated off the ground and features a long, sharp, and sickle-shaped ungual phalanx (claw). This particular trait, while now iconic, was not fully understood until the 1970s. The discovery of Deinonychus played a critical role in shifting our perception of dromaeosaurids, and more generally, theropods, anatomy, functional morphology, physiology, and behaviour. American palaeontologist John Ostrom’s (1969) description of Deinonychus highlighted the potentially lethal function of these sickle claws as possible weapons capable of tearing and disembowelling prey (Ostrom, 1969). However, more recent research suggests that this pedal ungual claw may have been primarily used to grasp and immobilise prey rather than for slashing and tearing (Carpenter, 1998; Norell & Makovicky, 1999; Manning et al., 2009; Fowler et al., 2009). This claw, which could grow over 6.5 cm in total length along the dorsal perimeter in adult Velociraptor specimens, remains one of the most distinctive and fascinating features of this dinosaur.
Over the past fifty years, palaeontological expeditions in Mongolia have yielded numerous Velociraptor specimens, including several nearly complete and fully articulated skeletons (Tab. 1). The most remarkable discovery is the famous “fighting dinosaurs” fossil, which captures a Velociraptor and a Protoceratops in a mortal combat. Discovered in 1971 by a Polish-Mongolian expedition at the Tugrik locality near the Flaming Cliffs, this fossil provides a rare and vivid snapshot of interspecies interaction (Kielan-Jaworowska & Barsbold, 1972). The two dinosaurs appear to have been buried alive in the midst of combat, possibly due to a sudden sandstorm or the collapse of a rain-soaked dune (Norell & Makovicky, 2004). The Velociraptor specimen is seen plunging its right sickle claw into the neck and the left into the belly of the Protoceratops, while the latter clamped the former’s right arm with its powerful beak. This interaction, once interpreted to having occurred at twilight, based on the finding that Velociraptor was nocturnal (Schmitz & Motani, 2011). However, re-evaluation of the correlation between scleral ring size and diel patterns are inconclusive for Velociraptor, making it impossible to ascertain the exact time of day of the encounter (Choiniere et al., 2021).
| Taxon | Specimen | Brief description | Formation | Locality | First referenced |
|---|---|---|---|---|---|
| Velociraptor mongoliensis | AMNH 6515 | a crushed but complete skull, associated with one of the raptorial second toe claws | Djadokhta Formation | Tugrikin-Shire, Omnogov, Mongolia | Osborn, 1924 |
| FPDM-V7220 | A complete skull | “ | “ | Azuma, 2005 | |
| MPC-D 100/24 | Almost complete skull, fragmentary postcranial elements | “ | “ | Barsbold, 1983 | |
| MPC-D 100/25 | Complete skeleton belonging to the “Fighting Dinosaurs” couple | “ | “ | Kielan-Jaworowska & Barsbold, 1972 | |
| MPC-D 100/54 | Almost complete specimen, associated with pterosaur bones (stomach content); one palaeopathological broken rib | “ | “ | Hone et al., 2012 | |
| MPC-D 100/976 | Partial skeleton and skull; punctures of Velociraptor teeth on the frontal bones | “ | “ | Norell et al., 1999; Norrell et al., 1995; Molnar, 2001 | |
| MPC-D 100/985 | Incomplete skeleton including fragmentary postcranial remains; bones scavenged by burrowing arthropods | “ | “ | Norell & Makovicky, 1997; Norell et al., 1997 | |
| MPC-D 100/1252 | Partial skull | “ | “ | Turner et al., 2007 | |
| MPC-D 100/2000 | A complete juvenile specimen | “ | “ | Barsbold & Osmolska, 1999; Tada et al., 2023 | |
| PIN 3143/8 | Almost complete skull with left mandibular ramus, lacking right temporal region; tip damaged | “ | “ | Barsbold & Osmolska, 1999 | |
| MPC-D 100/986 | Partial skeleton and fragmentary skull | “ | Chimney Buttes, Omnogov, Mongolia | Norell & Makovicky, 1997; Norell et al., 1999 | |
| Velociraptor sp. | MPC-D 100/982 | Almost complete skeleton, with a well-preserved skull and torso | “ | Bayn Dzak (“Volcano sublocality”), Omnogov, Mongolia | Norell & Makovicky, 1997; Norell & Makovicky, 1999 |
| ‘Velociraptor’ osmolskae | IMM 99NMBYM-3/3 | Both maxillae and left lacrimal | Bayan Mandahu Formation | Quarry SBDE 99BM-III, Urad Rear Banner, Inner Mongolia, China | Godefroit et al., 2008 |
| ?Velociraptor | MPC-D 100/981 | Fragmentary skeleton; ulna with quill knobs | Djadokhta Formation | Gilvent Wash, Omnogov, Mongolia | Turner et al., 2007 |
The discovery of the “fighting dinosaurs” brought Velociraptor to public attention almost fifty years after its initial discovery. The first reconstruction of Velociraptor depicting the battle with Protoceratops appeared in a 1975 book illustrated by Giovanni Caselli (Halstead, 1975). This book showcased many new and previously overlooked taxa, including the first-life reconstruction of Spinosaurus. However, Caselli’s depiction did not include the characteristic sickle claw on Velociraptor’s foot. The same year, the renowned palaeoartist Zdenek Burian also illustrated Velociraptor, portraying it in a dynamic pose but with a head shape more aligned with that of Deinonychus, a mistake later replicated by the Jurassic Park franchise. Throughout the 1980s, more accurate reconstructions began to emerge, thanks to the diligent efforts of palaeoartists such as Mark Hallett (Waldrop & Loomis, 1984) and Gregory Paul (Paul, 1988). These artists, along with Bakker (1986) were among the first to envision Velociraptor as a feathered dinosaur, anticipating the discovery of feathered dromaeosaurids in China in the following decades.
Paul’s (1988) work, including his book ‘Predatory Dinosaurs of the World’, suggested that Deinonychus and Velociraptor might be congeneric, with Deinonychus considered a junior synonym, though it was not based on detailed character analysis. Although this hypothesis was not widely accepted within the scientific community, it influenced Michael Crichton’s novel ‘Jurassic Park’, published in 1990. As a result, Velociraptor made its pop-culture debut portrayed as Deinonychus, an animal nearly twice its actual size. In reality, Velociraptor was closer in size to a wild turkey (Paul, 2017). The 1993 film adaptation by Steven Spielberg catapulted Velociraptor to global fame, depicting it as a highly intelligent, pack-hunting predator.
Despite the portrayal of Velociraptor in Jurassic Park, significant palaeontological discoveries in the late 20th and early 21st centuries have transformed our understanding of this dinosaur. Direct evidence of feathers in dromaeosaurids emerged in 1999 with the discovery of Sinornithosaurus (Xu et al., 1999). While no Velociraptor specimens preserve feathers, the presence of wing-like feather arrangements on the forelimbs of dromaeosaurids such as Zhenyuanlong and Microraptor, coupled with quill knobs on the ulna of a specimen referred to Velociraptor provides compelling evidence that it was also feathered (Turner et al., 2007). Although these feathers were unlikely to have enabled flight due to limb proportions and body mass, except in small-bodied species like Microraptor and Rahonavis, they might have played roles in reproduction, such as egg protection (Hopp & Orsen, 2004), or in courtship displays and other forms of display and/or communication (e.g., Zhang et al., 2010; Zelenitsky et al., 2012; Li et al., 2012).
Despite the cultural imprint left by Jurassic Park, the reality of Velociraptor as a feathered dinosaur remains underrepresented in the public imagination. Nonetheless, the film franchise has helped Velociraptor to become one of the most recognised, albeit misunderstood, dinosaurs. One hundred years after its description, advances in palaeontology have revealed Velociraptor as a small, feathered theropod with formidable sickle claws for restraining prey, jaws filled with recurved and serrated teeth, and neural anatomy indicative of a fast predator capable of tracking moving objects and hearing a wide range of frequencies (King et al., 2020). Even though direct evidence of feathers and pack-hunting remains elusive, ongoing research promises to uncover new aspects of Velociraptor biology.
A century since its first description, Velociraptor continues to be a focal point in palaeontological research. Our understanding of this dinosaur has grown substantially through new fossil discoveries, technological advancements, and refined analytical techniques. Recent studies have examined various facets of Velociraptor anatomy, behaviour, and ecology, offering a more nuanced view of this dynamic predator.
The Gobi Desert, particularly the Djadokhta Formation, remains a rich source of Velociraptor fossils. Among the most significant finds are nearly complete skeletons that have provided critical insights into the dinosaur’s anatomy and biology. Detailed examinations of the Velociraptor skull have revealed specialised features such as an enlarged olfactory bulb, suggesting a keen sense of smell that likely aided in hunting (Barsbold & Osmólska, 1999; Norell & Makovicky, 2004). Additionally, the unique structure of the braincase indicates advanced sensory processing capabilities, contributing to the predatory effectiveness (King et al., 2020).
Modern imaging techniques such as CT scans and 3D modelling, have allowed researchers to explore the internal structures of Velociraptor, uncovering details about its brain, sensory organs, and musculoskeletal system that were previously inaccessible. These technological advancements have enabled scientists to simulate movements and feeding behaviours in Velociraptor with unprecedented accuracy, providing deeper insights into its ecology and lifestyle.
In this contribution, to celebrate the 100th anniversary of the first description of Velociraptor, we present a comprehensive review of the current state of knowledge on this iconic non-avian theropod by discussing taxonomic and phylogenetic aspects, palaeobiological characteristics, the evolution of its iconography over time, and its influence on popular culture.
MATERIAL AND METHODS
The present state of the art on the various aspects of anatomy, general biology, feeding behaviour, metabolism, locomotion, palaeoneurology, palaeopathology, palaeoenvironment of life and iconographic evolution over time of Velociraptor was conducted on the basis of an extensive review of the existing literature, consisting of over 190 scientific publications (i.e., articles, book chapters, books; see References).
The phylogeny of Dromaeosauridae and phylogenetic position of Velociraptor was discussed based on the results of a cladistic analysis conducted on the data matrix by Wang et al. (2022) using the software PAUP* 4.0. A heuristic search was used as search option; this was run with 100 trees set as the number of maximum trees in memory (automatically increased by 100 if this limit was hit), 10000 random replicates, and the Tree Bisection Reconnection (TBR) as swapping algorithm. 252 most parsimonious trees (MPTs) of score 2151 were found. A strict and majority rule consensus tree (50% of frequency required) of the MPTs were then calculated and outputted. Such trees resulted in similar topologies, with the only exception being Saurornitholestinae, for which the OTUs were recovered as a single polytomy in the strict consensus tree. A similar analysis was run using the Czepiński (2023) data matrix, which is based on that of Powers et al. (2021a). 317 MPTs of score 406 were found. A majority rule consensus tree (50%) of the MPTs was then calculated and outputted. The two data matrices were chosen since they represent the recent outputs of different research groups; the aim of the analysis was to compare the resulting topologies obtained under the same standardised methodology; results of these analyses are briefly discussed in a dedicated section. For both analyses, consensus tree length and a diagnostic for each character are available as supplementary file (Appendices 2, 3).
In the section dedicated to the evolution of Velociraptor iconography over time, we also propose an updated reconstruction of Velociraptor mongoliensis. The reconstruction of Velociraptor provided in the section “Velociraptor iconography” is based on the specimen MUST SN1140/BM, digitised through photogrammetry following the procedure applied in several recent works on terrestrial tetrapod skeletons (e.g., Romano & Rubidge, 2021; Romano et al., 2021a, 2021b, 2022a, 2022b, 2023, 2024; Van den Brandt et al., 2024). The 3D model was reconstructed from 186 photos taken all around the skeleton using a 24-megapixel Canon EOS 750D (18 mm focal length). The skeletal model in 3D served as the base to sculpt the animal in flesh and bones. Upon the flesh, feathers and down were placed to fit the current knowledge and available data on related taxa. For the realisation of the in-vivo portrait, the digital sculpting program Maxon Zbrush 2023 was used. The program allows the insertion of background visual references, using multiple perspective views as a reference for sculpting, in combination with photogrammetry from the cast. Details such as scales and skin folds were added to the model manually. The plumage was reconstructed using the Fibermesh tool, which handles the realisation of filaments. The model was finally coloured using ZBrush digital painting tools.
A review of the systematic palaeontology of Velociraptorinae is provided as supplementary material as Appendix 1.
Known specimens of Velociraptor
The first Velociraptor specimen known to science is a complete yet mediolaterally crushed skull associated with an articulated ungual and penultimate phalanx of manual digit III (AMNH 6515) (Osborn, 1924) (Fig. 1). After WWII, while Cold War restrictions prevented North American teams from entering to communist Mongolia, expeditions by Soviet and Polish scientists, working with Mongolian colleagues, collected numerous dinosaurian remains, including specimens of Velociraptor. The most famous of these findings is the “fighting dinosaurs” (MPC-D 100/25) discovered on the 3rd of August 1971 (Kielan-Jaworowska & Barsbold, 1972; Barsbold, 1974). Other recovered Velociraptor specimens include MPC-D 100/24, an incomplete skeleton with fragmentary postcranial elements and an almost complete skull (Barsbold, 1983; Barsbold & Osmólska, 1999), MPC-D 100/2000, a juvenile complete skeleton (Barsbold & Osmólska, 1999), and PIN 3143/8, an almost complete skull (Barsbold & Osmólska, 1999).
In the early 1990’s several joint expeditions of the Mongolian Academy of Sciences and the American Museum of Natural History brought to light more dinosaurian skeletons, including new abundant Velociraptor material (e.g., Norell et al., 1995; Norell & Makovicky, 1997, 1999; Norell et al., 1997). Table 1 includes most of the published specimens belonging to this genus.
Specimen MPC-D 100/982 was collected in 1992 at Bayn Dzak locality (“Volcano sublocality”), not far from Tugrikin-Shire locality. Although this specimen was originally referred to as V. mongoliensis, it shows clear differences in the pelvic region and neuroanatomy (Kundrát, 2004; Norell & Makovicky, 1999). A recent analysis suggests it might represent a different and as-yetunnamed species (Powers et al., 2020; Ruebenstahl et al., 2023), and therefore here referred to Velociraptor sp. (Fig. 2).
- Right lateral side (a) and left lateral side (b) of MPC-D 100/982.
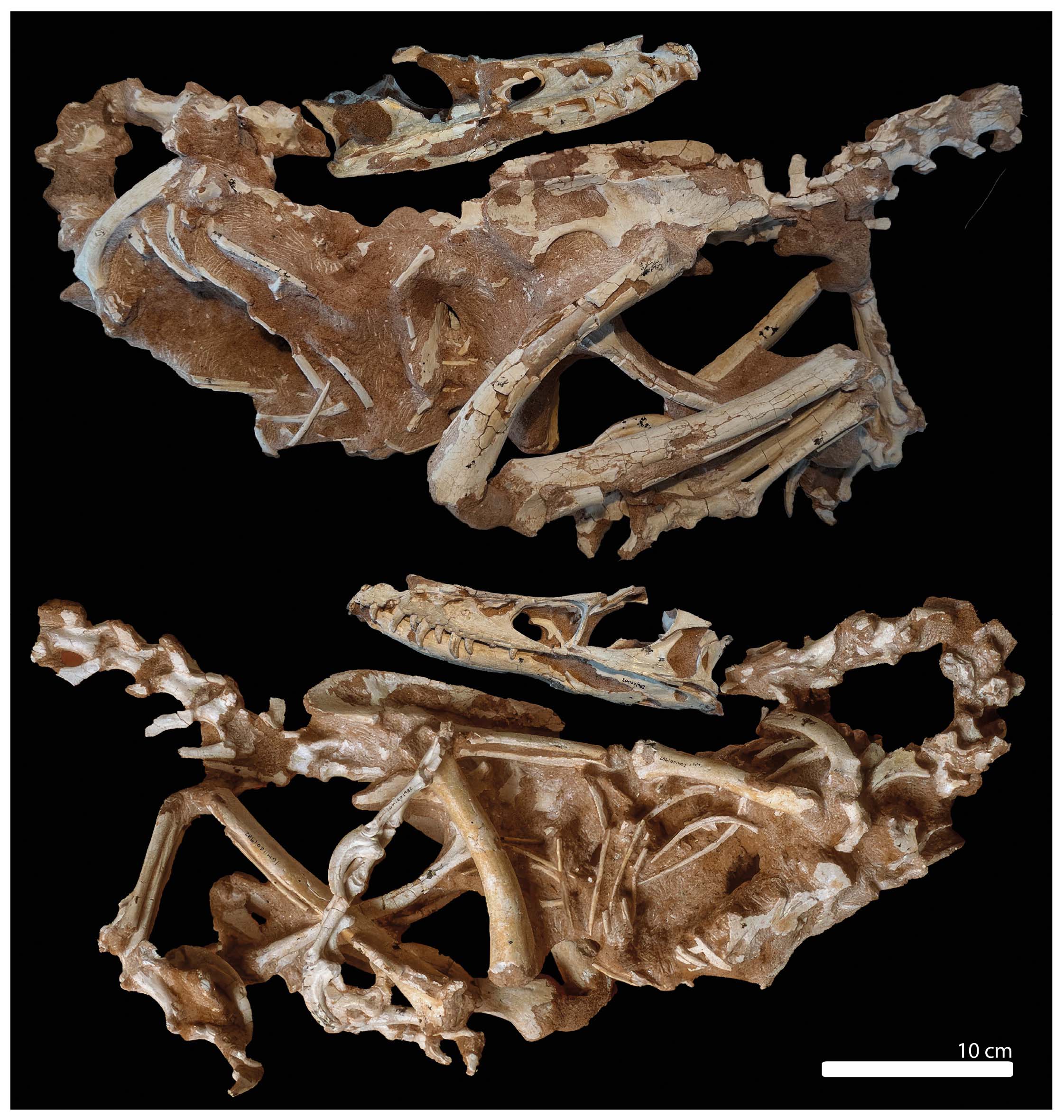
Specimen IMM99NM-BYM-3/3 was recovered during a Sino-Belgian expedition in 1999 and later named Velociraptor osmolskae by Godefroit et al. (2008). However, phylogenetic analyses such as Evans et al. (2013) (but see also Czepiński, 2023) found it more closely related to Linheraptor than to Velociraptor, therefore suggesting that this species may not belong to the genus Velociraptor. Since that time, the specimen has been referred to ‘Velociraptor’ osmolskae, waiting for the erection of a new genus.
Specimen MPC-D 100/981, a fragmentary skeleton including an ulna with quill knobs, was originally referred to V. mongoliensis (Turner et al., 2007) (Fig. 3). Nonetheless, its fragmentary condition does not support such specific attribution, and the specimen is preferably referred to Velociraptorinae indet. or ?Velociraptor (Napoli et al., 2021).
- (a) Dorsal view of right ulna of Velociraptor MPC-D 100/981. (b) Detail of the white box in (a) showing six evenly spaced feather quill knobs highlighted in the line drawing in (c). (a) and (b) are modified from Turner et al. (2007).

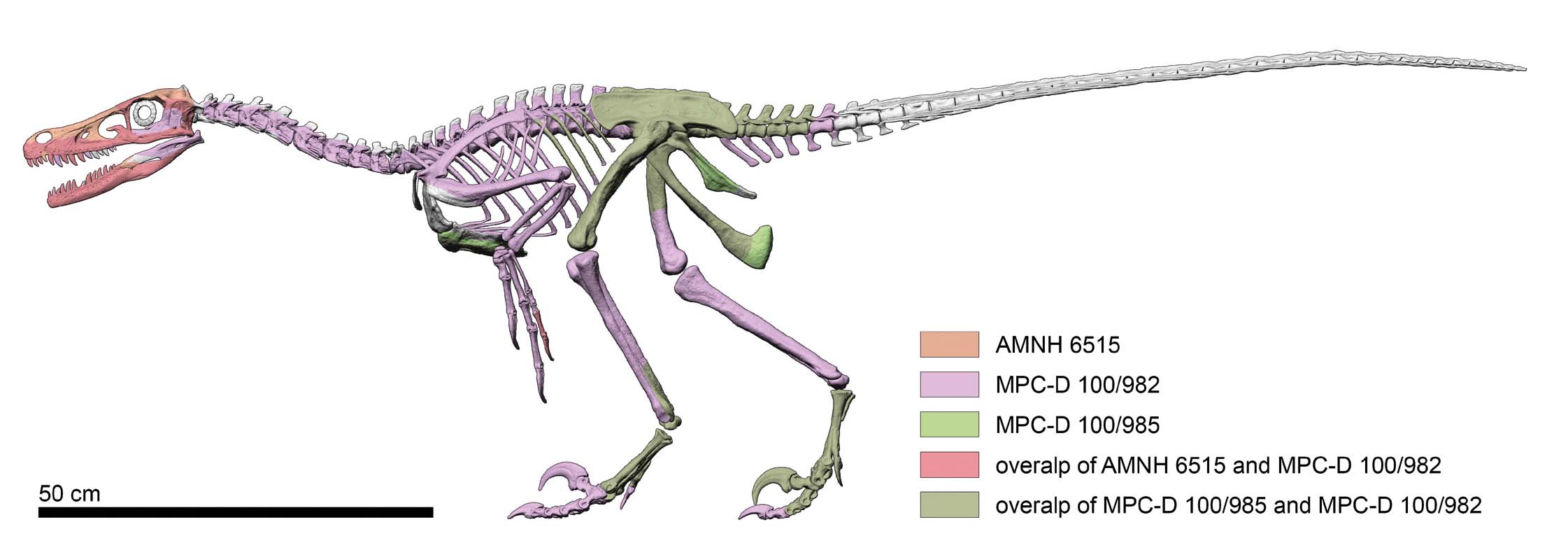
Figure 4 shows an updated skeletal reconstruction of Velociraptor, including available material of some selected specimens.
- Skeletal reconstruction of Velociraptor. Colour schematic representation of the available material belonging to AMNH 6515, MCP-D 100/982, and MCP-D 100/985. Size is based on MCP-D 100/25.
TAXONOMY OF EUDROMAEOSAURIA AND DEFINITION OF VELOCIRAPTORINAE
Sereno (1998) defined the group Velociraptorinae Barsbold 1983 as all dromaeosaurids more closely related to Velociraptor than to Dromaeosaurus, which along with its purported sister taxon Dromaeosaurinae Matthew & Brown, 1922 constituted the clade Dromaeosauridae (Matthew & Brown, 1922) Russell 1969, before the recognition of other taxa such as Microraptorinae and Unenlagiinae.
Dromaeosaurinae typically includes at least three species: Utahraptor ostrommaysi, Achillobator giganticus, and Dromaeosaurus albertensis (Senter et al., 2012; DePalma et al., 2015; Napoli et al., 2021; Powers et al., 2021a; Wang et al., 2022); nonetheless, recent analyses, recovered Achillobator and Utahraptor as Velociraptorinae (Currie & Evans, 2020; Jasinski et al., 2020) whereas Deinonychus is either recovered as a member of Velociraptorinae or Dromaeosaurinae (Napoli et al., 2021; Powers et al., 2021a; Wang et al., 2022).
The clade Saurornitholestinae was recognised for the first time by Longrich & Currie (2009) and recovered as the sister taxon to a clade formed by Dromaeosaurinae and Velociraptorinae. It typically includes three species: Atrociraptor marshalli, Bambiraptor feinbergi, and Saurornitholestes langstoni (Longrich & Currie, 2009; Jasinski et al., 2020). Bambiraptor feinbergi has recently been suggested to be a junior synonym of Saurornitholestes langstoni (Pittman et al., 2020), of which it may represent a juvenile individual; Shri, Kuru, and Acheroraptor are either recovered as Saurornitholestinae or Velociraptorinae (Napoli et al., 2021; Turner et al., 2021; Powers et al., 2021a; Wang et al., 2022; Czepiński, 2023).
The three clades Velociraptorinae, Dromaeosaurinae, and Saurornitholestinae belong to Eudromaeosauria, first recovered and defined by Longrich & Currie (2009) as the most inclusive clade containing Dromaeosaurus, Velociraptor, Deinonychus, and Saurornitholestes, their most recent common ancestors and all its descendants. Dromaeosaurinae and Velociraptorinae are often recovered as sister clade, with Saurornitholestinae forming a basally branching radiation within Eudromaeosauria (Longrich & Currie, 2009; Evans et al., 2013; Powers et al., 2020; Napoli et al., 2021; Turner et al., 2021); conversely, in other analyses, Velociraptorinae is sister to Saurornitholestinae, with Dromaeosaurinae being a basal clade of Eudromaeosauria (Powers et al., 2021a; Wang et al., 2022).
Systematic Palaeontology
Dinosauria Owen 1842
Theropoda Marsh 1881
Dromaeosauridae Matthew and Brown 1922
Velociraptorinae Barsbold 1983
Genus Velociraptor Osborn 1924
Type species: Velociraptor mongoliensis Osborn 1924
Velociraptor mongoliensis Osborn 1924
Referred specimens: see Table 1.
Type locality: Bayn Zag (“Flaming Cliffs”), Ömnögov, Mongolia.
Type horizon: Djadokhta Formation, Campanian, Upper Cretaceous.
‘Velociraptor’ osmolskae Godefroit et al. 2008
Type locality: quarry SBDE 99BM–III, Bayan Mandahu, Mongolia.
Type horizon: Bayan Mandahu Formation, Campanian, Upper Cretaceous.
Remarks: distinct from V. mongoliensis by the size and shape of the promaxillary fenestra and the elongate ratio of the rostral plate of the maxilla. Because it is not recovered as the sister taxon of Velociraptor mongoliensis in some recent analyses (e.g., Powers et al., 2021a; Czepiński, 2023; but see Wang et al., 2022), this species is here refered to as ‘Velociraptor’ mongoliensis. The phylogenetic placement (or affinity) of this species has yet to be confirmed but the general consensus suggests that this taxon should be referred to a distinct genus.
Velociraptor sp. Powers 2020
Type locality: Volcano sub-locality, Flaming Cliffs, Bayan Dzak, Mongolia.
Type horizon: Djadokhta Formation, Campanian, Upper Cretaceous.
Remarks: MPC-D 100/982, the single specimen included in this as-yet-unnamed taxon, was initially tentatively assigned to V. mongoliensis. Nonetheless, since its discovery, differences between it and V. mongoliensis have been identified in the anatomy of the pelvic region and the neurocranium (Norell & Makovicky, 1997, 1999; Kundrát, 2004). Powers (2020) confirmed that this specimen represents a third species of Velociraptor, which he named Velociraptor vadarostrum and provided a character-based diagnosis. Because the naming of this species was in a M.Sc. thesis (Powers, 2020) rather than in a peer-reviewed journal, the binomen must be considered a nomen nudum. Powers et al. (2020, 2021a, 2021b, 2021c) and Ruebenstahl et al. (2023) confirmed the existence of this third species, which to date remains unnamed.
RESULTS OF THE PHYLOGENETIC ANALYSIS
Results of the cladistic analyses performed on the two datamatrices (A and B, see Material and Methods section) reveal that Dromaeosauridae includes Halszkaraptorinae, Unenlagiinae, Microraptorinae, Yixian dromaeosaurids, and Eudromaeosauria. The clade Averaptora (Troodontidae+Avialae) is found only in A; in B, Avialae is sister to Troodontidae+Dromaeosauridae (Fig. 5). According to analysis A, Balaur belongs to Avialae, and is found to be sister to Aves, consistently with Cau et al. (2015) contra previous hypotheses (see above for its systematic). In A, clade Halszkaraptorinae+Unenlagiinae is found as sister of Shanag+more derived Dromaeosauridae, whereas in B Halszkaraptorinae is recovered as the basalmost member of Dromaeosauridae. On the other hand, Unenlangiinae in A is sister to Halszkaraptorinae, whereas in B it occupies a polytomy with Yixiang dromaeosaurids more derived than Microraptorine, but more basal than Eudromaeosauria. In A, Shanag is recovered more basal than Microraptorinae, contra the original analysis of Wang et al. (2022) where it is sister to Yixian dromaeosaurids + Eudromaeosauria.
- Cladograms (50% majority rule consensus trees) obtained with Wang et al. (2022) (a) and Czepiński (2023) (b) data matrix. The two results represent different outputs of different research groups and are here presented to comment on the differences in the topology obtained. See text for details on method and discussion. Abbreviations: P=Paraves; A=Averaptora; D=Dromaeosauridae. 0-100 numbers indicate frequency of occurrence for each node. Silhouettes used in the Figure are available at phylopic.org. Shared under public domain: Archaeopteryx lithographica by T. Michael Keesey; Sinovenator changii by Tasman Dixon. Shared under CC BY-SA 3.0 (creativecommons.org/licenses/by-sa/3.0/): Atrociraptor marshalli, Balaur bondoc, Deinonychus antirrhopus, and Sinornithosaurus millenii by Emily Willoughby; Mahakala omnogovae by Scott Reid; Tianyuraptor ostromi by Mette Aumala; Saurornitholestes langstoni and Velociraptor mongoliensis by Scott Hartman. Shared under CC BY 4.0 (creativecommons.org/licenses/by/4.0/): Buitreraptor gonzalezorum by Ivan Iofrida.
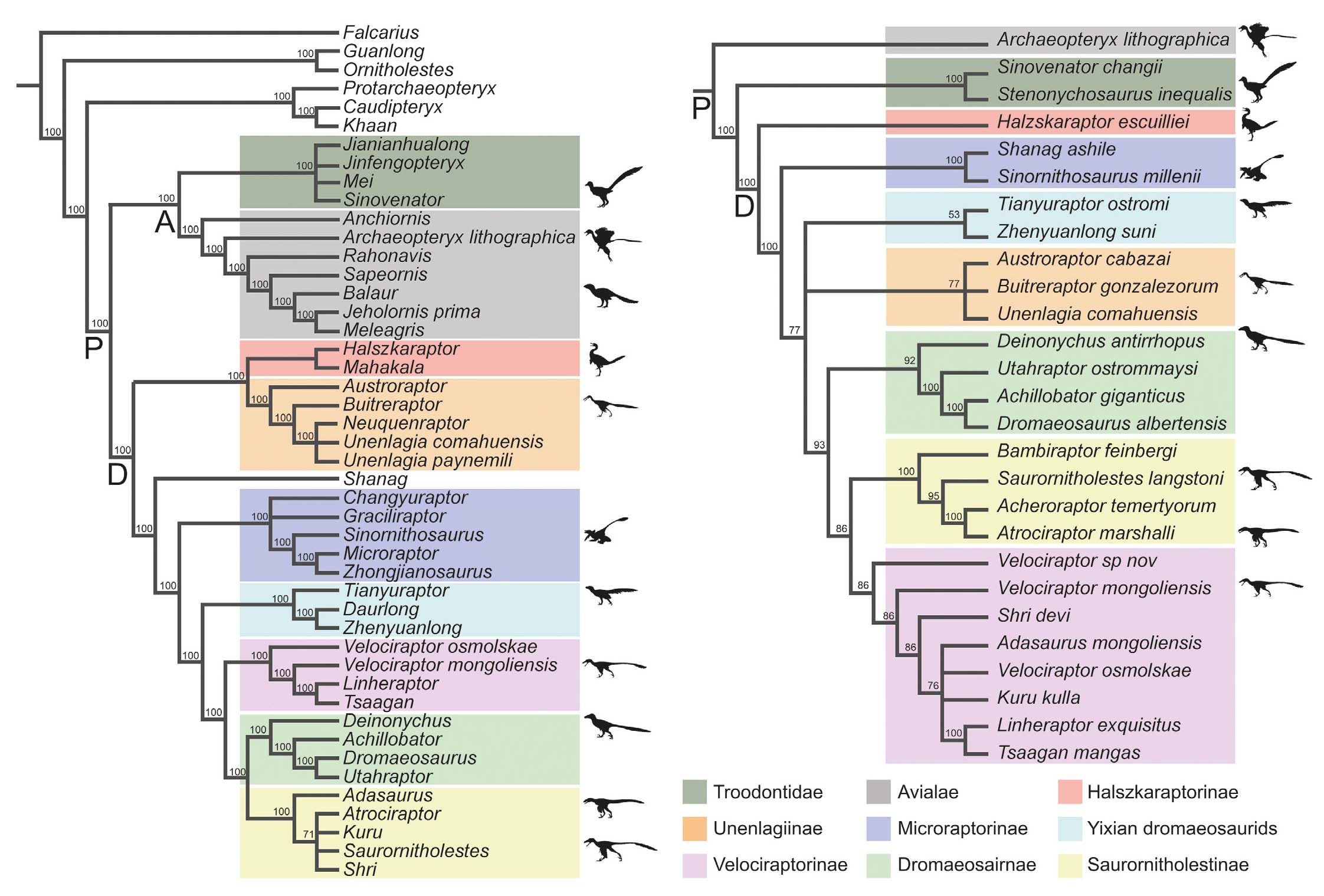
In both analyses, Eudromaeosauria comprises Velociraptorinae, Dromaeosaurinae, and Saurornitholestinae. In A, Velociraptorinae is sister to Saurornitholestinae+Dromaeosaurinae, whereas in B Dromaeosaurinae is sister to Saurornitholestinae + Velociraptorinae. In B, Velociraptorinae comprises Velociraptor, Linheraptor, Tsaagan, Shri, Adasaurus, and Kuru, whereas in A, the latter three genera belong to Saurornitholestinae. Likewise, in B Adasaurus is no longer recovered as the basalmost member of Eudromaeosauria, contra Powers et al. (2021a: Fig. 18A) and Czepiński (2023: Fig. 6A).
- Dorsolateral view of MPC-D 100/986 tail.

None of the two consensus trees resulting from analyses A and B recovered “Velociraptor” osmolskae in a monophyletic clade with Velociraptor mongoliensis and/or Velociraptor sp. Moreover, despite previous results (e.g., Powers et al., 2020; 2021a: Fig. 18A), in B Velociraptor sp. is not recovered sister to Velociraptor mongoliensis, but is instead classified as the basalmost member of the clade Velociraptorinae. This result may be the consequence of two reasons. First, the poorly resolved topology of Eudromaeosauria. Secondarily, under an overly reductive interpretation, taxa such as Tsaagan and Linheraptor could be considered junior synonyms of Velociraptor, which would be the generic affiliation for all Velociraptorinae. Accepting the latter explanation would allow considering “Velociraptor” osmolskae to belong to Velociraptor. Nonetheless, this solution cannot be recommended, given our current knowledge of velociraptorine taxonomy, both because the topology of Eudromaeosauria is not well-resolved and because of the presence of clear character distinctions between the mentioned taxa. In this context, we once again support the proposal to establish a new genus for “Velociraptor” osmolskae, and to clearly redefine the specimen MPC-D 100/982 to assess its systematic position and, ideally, advance the resolution of Velociraptorinae phylogeny.
DIAGNOSIS OF VELOCIRAPTOR
A notable gap in our current understanding of Velociraptor is the absence of an up-to-date, comprehensive diagnosis. There is no published updated formal list of autapomorphic traits or differential diagnostic features that reliably distinguish Velociraptor from other closely related taxa. Furthermore, recent phylogenetic analyses have failed to identify a consistent list of diagnostic traits for the genus Velociraptor. The comparison of two major parsimony-based analyses (Wang et al., 2022; Czepiński, 2023; see above) illustrates the complexity of the issue. In the analysis by Czepiński et al. (2023), only a single autapomorphic trait optimised as diagnostic for Velociraptor in all MPTs—the size of the denticles on its teeth (character 85). This contrasts with the findings of Wang et al. (2022) who identified 12 diagnostic characters, exclusively related to the cranium, though one related to the dentition and has been miscored (absence of 4th premaxillary alveolus); in fact, Barsbold & Osmolska (1999) and Currie & Evans (2020) list four premaxillary teeth in Velociraptor. Even if a detailed analysis is beyond the scope of this review, a brief discussion of the challenges involved in defining these diagnostic traits is warranted.
The well-known arched nasal bone is one of the most peculiar characters historically associated with Velociraptor. While this trait is observed in the holotype and several other specimens, including MPD-C 100/24, 100/25 and 100/986, it is absent in Deinonychus (Brownstein, 2021) and taxa referred to saurornitholestines (Currie & Evans, 2020). However, the nasals are absent in several relevant taxa, including “V.” osmolskae, Adasaurus and Shri, making uncertain the autapomorphic status of this trait.
The lack of overlap between the recovered topologies in the phylogenetic analyses published in the literature (e.g., Wang et al., 2022; Czepiński, 2023) highlights the ongoing uncertainty and a missing consensus on the diagnosis of this genus. This inconsistency raises questions about whether the current diagnostic framework is sufficient to capture the morphological variation within the taxon.
Future work should aim to resolve these discrepancies by integrating new fossil descriptions (such as MPC-D 100/982) with updated phylogenetic analyses. Until such progress is made, any discussion of Velociraptor’s diagnostic characters remains tentative.
General anatomy
Velociraptor had a slender, agile body with a length of up to two metres and stood around 1 m tall at the hip (Kielan-Jaworowska & Barsbold, 1972; Norell & Makovicky, 1997; Norell & Makovicky, 1999). The skull of Velociraptor was particularly elongated (Fig. 3) with a slender snout bearing sharp, ziphodont teeth (Barsbold & Osmólska, 1999). This animal is characterised by using only two toes to walk and by possessing a raised sickle-shaped ungual phalanx, as seen in other basal paravian (Li et al., 2007).
Skull
The following general (but not exhaustive) description of the Velociraptor skull is based on AMNH 6515, MPC-D 100/25, MPC-D 100/54, MPC-D 100/982, and IMM 99NM-BYM-3/3. The skull is characterised by its elongated and slender shape. The nasals display a peculiar dorsally concave curvature that is unique among dromaeosaurid taxa that preserve the nasal as discussed above. The antorbital region occupies approximately two-thirds of the entire skull length, which is a typical ratio among predatory theropods. The premaxilla in Velociraptor mongoliensis is longer than tall, whereas the opposite condition is observed in the Velociraptor specimen MPC-D 100/982 (Fig. 2). The narial opening is oval and anteroposteriorly longer than taller; anterior to it, a small prenarial fossa pointing ventrally is present. Posterodorsally, the premaxilla contacts the nasals, whereas ventrally it articulates with the maxilla through an extremely elongated subnarial process that reaches far posterior to the naris. The maxilla is triangular in shape, with a concave posterior margin defined by the anterior arc of the antorbital fenestra. The antorbital fossa also houses at least two more antorbital openings (the maxillary fenestra and 1-2 promaxillary foramina), of which the smallest one reaches the anterior tip of the antorbital fossa. The maxillary fenestra is centred on the antorbital fossa and well-removed from its ventral edge. The maxilla bears 11 tooth sockets. Maxilla and premaxilla proportions have been used recently to shed light on the systematics of Eudromaeosauria. In Velociraptor the antorbital fenestra and the maxilla are more elongated than in other dromaeosaurids (e.g., Powers et al., 2021a; Czepiński, 2023). The presence of a prefrontal remains unknown. The nasals are located dorsal to the maxilla and contact anteroventrally the premaxilla, with which they form the external naris opening, whereas posteroventrally they contact the dorsal ramus of the maxilla. Posteriorly, the nasal contacts the frontal, and posterolaterally the lacrimal. The lacrimal is roughly T-shaped; the ventral ramus is straight and thin in lateral view and is incised by the antorbital fossa rostrally. Its ventralmost portion flares to assume a triangular shape, which contacts the jugal. The junction of the three rami forms a low, laterally projecting lacrimal crest. The lacrimal forms the posterodorsal margin of the antorbital fenestra and the anterodorsal margin of the orbital fenestra. The frontal is four times longer than wide, as well as four times longer than the parietals, with which it contacts posteriorly. The parietals are fused and have a complex three-dimensional shape; they form the posteromedial quarter of the upper temporal fenestra. They bear a low midline sagittal crest, as is typical for dromaeosaurids. The upper temporal fenestra is roughly rounded, with its posterolateral margin defined by the squamosal and its anterior margin defined by a sigmoidal ridge extending from the postorbital and across the dorsal surface of each parietal (Currie, 1995). The lower temporal fenestra is roughly reniform in shape, its dorsal margins are formed by the postorbital and the squamosal anteriorly and posteriorly, respectively, and its ventral margin by the jugal and quadratojugal (respectively anteriorly and posteriorly). The jugal is roughly triangular, with the dorsal ramus contacting the descending process of the postorbital anteroventrally, the anterior ramus contacting the posterior ramus of the maxilla, and the posterior ramus contacting the anterior ramus of the quadratojugal laterally. The quadratojugal also has three rami and has an inverted ‘T’ shape like in Deinonychus (Ostrom, 1969) and Tsagaan (Norell et al., 2006). The dorsal ramus contacts the descending prequadratic process of the squamosal and the lateral process of the quadrate shaft, the medial side of the ventral ramus contacts the jugal, and the posterior process contacts the lateral side of the quadrate condyles. The angle between the dorsal and posterior rami forms the border of an enlarged quadrate foramen characteristic of dromaeosaurids.
The braincase is poorly pneumatised compared to other paravians, especially in derived troodontids. The anterior tympanic recess forms a small pocket posterior to the exit for cranial nerve V and ventral to that of cranial nerve VII (Norell & Makovicky, 2004). The dorsal tympanic recess presents as a deep pocket opening dorsolaterally near the base of the paroccipital process, whereas the posterior tympanic recess forms a pocket adjacent to the middle ear opening. Ventrally, the basisphenoid is perforated by a pair of foramina leading into the basisphenoid recess.
The dentary is thin and elongated, with the anteriormost portion slightly curved dorsally; it possesses 14-15 tooth sockets. At its posterior margin, the dentary articulates with the surangular and angular at the external mandibular fenestra, contributing only a small portion of its anterior margin. This fenestra is elongated, four times anteroposteriorly longer than dorsoventrally tall, oval, and its posterior margin is slightly tilted ventrally. More posteriorly, the surangular fenestra is relatively small and overhung by a prominent surangular shelf, as in many dromaeosaurids. The sizes, proportions, and orientation of these two openings can slightly vary from specimen to specimen. The splenial is exposed on the lateral side of the mandible, but the extension of this exposure shows some intraspecific variability as well. It cannot be excluded that this variability might also be caused by different taphonomical factors.
Regarding the dentition, the premaxilla is characterised by four teeth with a slight curvature, with the two anterior teeth being of the same size and approximately twice the size of the third and fourth teeth in specimen GIN 1000124 (Barsbold & Osmólska, 1999). The maxilla is characterised by 11 slender, posteriorly curved teeth, with every second tooth being longer than the adjacent ones, at least in the anterior half of the tooth row (Barsbold & Osmólska, 1999). The dentary in the holotype has a tooth count ranging from 14 to 15 (Barsbold & Osmólska, 1999).
The dentition of Velociraptor exhibits several distinctive dental traits characteristic of dromaeosaurids, particularly within Velociraptorinae. Following the review by Hendrickx et al. (2019) the teeth are ziphodont, featuring denticulated mesial and distal carinae in at least some lateral teeth, with a denticle size disparity index (DSDI, see Hendrickx et al., 2019) typically exceeding 1.2. Notably, in most lateral teeth, the mesial carina does not extend to the cervix, a feature observed in all velociraptorines except Tsaagan, whose mesial denticulate carina is absent. Additionally, some lateral teeth display a figure-of-eight-shaped cross-sectional outline at the base of the crown, a feature present in most velociraptorine taxa other than Deinonychus.
The mesial dentition of Velociraptor is characterised by the lack of serrations along the mesial carina, which is either entirely absent or smooth. The distal carina, however, remains denticulated, consistent with the condition observed in most theropod clades. Additionally, Velociraptor exhibits fluted labial surfaces on its mesialmost teeth, a feature also documented in Ceratosaurus and Scipionyx (Hendrickx et al., 2019). The number of flutes present on the first premaxillary tooth (Rpm1) in Velociraptor (AMNH 6515), ranges from three to six (Hendrickx et al., 2019).
In lateral teeth, Velociraptor conforms to the general pattern observed in Velociraptorinae, with denticulated mesial and distal carinae, albeit with the mesial carina failing to reach the cervix. Some lateral teeth display longitudinal ridges. The absence of apically hooked denticles differentiates Velociraptor from some other theropods, as it instead possesses symmetrically to asymmetrically convex serrations (Hendrickx et al., 2019).
Postcranium
The following description of the Velociraptor postcranium is based on MPC-D 100/25, MPC-D 100/54, MPC-D 100/976, MPC-D 100/2000, MPC-D 100/982, MPC-D 100/986, MPC-D 100/985, MPC-D 100/3503. Although the postcranium of Velociraptor is often used alongside that of Deinonychus as an exemplar of eudromaeosaurian anatomy, no autapomorphies have been identified in the literature, nor do any postcranial traits optimise as such in either of the two published phylogenetic analyses reviewed herein.
Vertebral column: the neck is strongly curved, powerfully built, and includes ten short and robust cervicals with wide neural arches and prominent epipohyses. Cervical ribs are short and are fuse to their respective vertebrae in some specimens (MPD-C 100/976, 100/986). Anterior trunk vertebrae bear large hypapophyses. More posterior trunk vertebrae have centra that are much taller than long and bear laterally projected parapophyses that extend out from their base for a distance comparable to the diameter of their articular surface (Norell & Makovicky, 1999). All presacral vertebrae save the last one or two have pneumatic foramina on their centra. The sacrum comprises five vertebrae in most specimens, though six are present in some larger and presumably older individuals. Sacrals exhibit partially fused neural spines.
The tail comprises around 25 vertebrae, and the transition point between proximal and distal caudals occurs around the seventh caudal vertebra. Extremely elongated prezygapophyses and stiffened tendons originate from the posteriormost available vertebra and more anterior elements, and reach proximally to the fourth caudal. These structures would have made the majority of the tail very rigid.
Pectoral girdle. It is composed of scapulocoracoids, the furcula (Fig. 7), and paired sternal plates. At least three ossified sternal ribs articulate with each sternal plate. The scapular blade is narrow and in lateral view widens slightly distal to the glenoid. The suture between the scapula and coracoid cannot be distinguished. Scapulocoracoids are arched all along their length and prominently flex at the base of the scapula. The coracoids are large, quadrangular, and flexed at the level of the coracoid tubercle, thus shifting the attached scapula onto the dorsal region of the ribcage. Each coracoid bears a large subglenoid shelf along the posterior edge. The furcula is thin, apneumatic, and its lateral rami are set at an obtuse angle to each other and are slightly arched along their lengths.
- Furcula of MPC-D 100/976 modified from Norrell et al. (1997) (a) and line-drawing of it (c).

Forelimb. The humerus is robust with a relatively long deltopectoral crest. The ulna is more robust than the radius, though not to the degree seen in e.g. Microraptor, and has a posteriorly curved shaft. Among dromaeosaurids, Velociraptor possessed a relatively large manus with three elongated digits, each culminating in a strongly curved ungual. Of the three digits, the second is the longest, whereas the first digit is the shortest, most robust, and bears the largest ungual. The arrangement of carpal bones prevented the pronation of the wrist, compelling the manus to be held with the palmar surface facing medially, close to the trunk, while in a resting position.
Pelvic girdle. The pelvic bones do not show pneumatic foramina. The ilium is dolichoiliac, and the preacetabular portion is shorter than the postacetabular one. In dorsal view, the upper border of each ilium exhibits a sigmoid profile, laterally concave anteriorly and convex posteriorly (Norell & Makovicky, 1997). One of the most striking features of the Velociraptor pelvis is the degree of pubic retroversion (155° between pubis and ilium). This degree of retroversion is more similar to Archaeopteryx lithographica than to Deinonychus antirrhopus (Ostrom, 1974). The ischia are about half as long as the pubes; they are not fused, but they contact each other distally and have flattened shafts with a rounded lateral ridge as in other dromaeosaurids; the ischial proximal end is anteroposteriorly expanded, making the bone T-shaped in lateral view.
Hindlimb. The femur is typically paravian in having a trochanteric crest, globular head offset by a distinct neck, and a prominent posterior trochanter and trochanteric shelf (Norell & Makovicky, 1999; Hutchinson, 2001). The fourth trochanter is variably developed, being reduced in most specimens but larger in at least one individual. The crus is marked by a slender fibula that reaches the ankle, a more robust tibia, and proximal tarsals in which the astragalus makes up ~80% of the distal articulation and bears a tall, triangular ascending process. The metatarsus is compact and relatively short compared to unenlagiines, microraptorines, and troodontids, but similar to other eudromaeosaurs like Deinonychus (Ostrom, 1969). Metatarsal III has a robust shaft that is well exposed in both extensor and plantar views. The foot features four digits, of which the first is reduced as in most theropods. The second digit bore a relatively large, sickle-shaped claw, a characteristic trait shared among dromaeosaurid and troodontid dinosaurs. This second digit, renowned for its unique anatomy in Velociraptor, was highly modified and held retracted above the ground, so the animal would have walked and run mostly only on digits III and IV (Barsbold, 1983; Norell & Makovicky, 1997).
Feathers
The study of feathers in non-avian dinosaurs has witnessed remarkable advancements in the last two decades, shedding light on the potential existence of feathered forelimbs in Velociraptorinae and plumage covering almost the entire body in some dromaeosaurids.
Velociraptor specimens, particularly those collected by the joint Mongolian Academy of Sciences-American Museum of Natural History (MAE-AMNH) expeditions, played a significant role in the debate on the origin of birds during the 1990s and early 2000s. Detractors of the hypothesis that birds are dinosaurs argued that the similarities between birds and theropod dinosaurs, including and especially Velociraptor, were either due to convergent evolution (Feduccia, 1999) or, in some cases, proposed even more radical ideas, such as dromaeosaurids being descended from birds (Feduccia et al., 2007). Papers such as Norell et al. (1997), which documented the presence of a furcula in Velociraptor mongoliensis MPC-D 100/976 (Fig. 6), played a critical role in reinforcing the theropod-bird link. This finding was met with strong opposition from critics like Feduccia & Martin (1998) thought for reasons based on unsupported assumptions. Similarly, debates surrounding the alleged ‘hypopubic cup’ of Archaeopteryx (Ruben et al., 1997; Norell & Makovicky, 1999) were also relevant in shaping the discussion.
Direct evidence of feathers in dromaeosaurids first came to light in 1999 with the discovery of Sinornithosaurus (Xu et al., 1999). The presence of winglike arrangements of pennaceous feathers on the forelimbs of dromaeosaurids Zhenyuanlong (Lü & Brusatte, 2015) and Microraptor (Xu et al., 2003), as well as in other outgroups, renders it parsimonious that they were also present in Velociraptor. Turner et al. (2007) identified small bumps on the posterior side of the ulnar shaft of a specimen referred to Velociraptor mongoliensis, which in terms of their number and distribution resemble the quill knobs that are attachment sites of wing feathers in birds. The fragmentary remains described by the authors are insufficient to support referral to this genus, although Napoli et al. (2021) suggest treating the specimen as an indeterminate velociraptorine. Turner et al. (2007) suggest that the quill knobs could possibly reflect the retention of feathers from smaller volant ancestors, but also have other functions. Thermoregulatory effects were probably insignificant if plumage is restricted to the ulna (Turner et al., 2021), but feathers could instead have been used with display functions (Norell & Xu, 2005), to regulate the temperatures of nests (Hopp & Orsen, 2004), or to complement locomotion during running and while hunting (Dial, 2003).
Jasinski et al. (2022) describe papillae on the ulnar ridge of Dineobellator notohesperus, considering them to be evidence that the animal had winglike plumage on its forelimbs. These authors state that while there have been theories that dromaeosaurid wings served as stabilizers during predatory attacks (Fowler et al., 2011) without acting as visual signals that draw the attention of predators or prey, feathers can serve as identifiers, species recognition markers, and/or sexual display elements (Jasinski et al., 2022). Modern raptors demonstrate that sophisticated colour patterns are still possible and can be used to both attract mates and disguise them as predators (e.g., Hill & McGraw, 2006; Holt et al., 1990; Snyder & Snyder, 2006).
The discussion of the potential multiple functions of dromaeosaurid feathers, as well as in any other feathered theropod, highlights the actual need for further investigations into their primary and secondary roles. Beyond the already mentioned ‘traditional’ hypotheses, feathers in Dromaeosauridae could speculatively have served as sensory organs akin to the filoplumes of extant birds, providing a heightened awareness of their surroundings, or being used to communicate through feather movement, such as inflating with feathers, as both during courtship or as a menacing symbol.
VELOCIRAPTOR PALAEOECOLOGY
Feeding habits
The diet of Velociraptor is relatively well known thanks to several direct pieces of evidence providing crucial insights into its feeding behaviour. The most famous example of direct evidence is the already mentioned “fighting dinosaurs” specimen from the Upper Cretaceous Djadokhta Formation of Mongolia (Kielan-Jaworowska & Barsbold, 1972; Holtz, 2003). As briefly mentioned above, this remarkable fossil preserves a Velociraptor locked in combat with a Protoceratops andrewsi, suggesting an active predation event. The dromaeosaurid’s sickle-shaped claw is positioned near the neck of the herbivore, implying a targeted attack strategy, while its left hand grips the frill of the Protoceratops. However, alternative interpretations exist, including the possibility that Velociraptor was acting in self-defence rather than actively hunting (Osmólska, 1993). Nonetheless, this specimen provides a rare instance of a theropod-prey interaction captured in the fossil record (Carpenter, 1998).
Further evidence supporting Velociraptor’s dietary habits derives from taphonomic analysis of bite marks and shed teeth. Fossil specimens from Bayan Mandahu, Inner Mongolia, include Protoceratops bones bearing feeding traces attributable to Velociraptor, suggesting that this theropod consumed the neoceratopsian either through active predation or scavenging (Hone et al., 2010). The observed bite marks, particularly those concentrated on the jaw region, indicate, according to the authors, late-stage carcass consumption, which may point toward scavenging rather than primary predation. However, given the prevalence of Protoceratops and Velociraptor in Djadokhtaequivalent formations, it is reasonable to infer that these interactions were relatively common (Hone et al., 2010).
In addition, the discovery of a pterosaur bone within the chest cavity of a Velociraptor provides direct evidence of ingestion, demonstrating that this dromaeosaurid incorporated a diverse range of prey into its diet (Hone et al., 2012).
Concerning indirect evidence of feeding behaviour, most discussions of dromaeosaurid lifestyle have focused on the enlarged raptorial second pedal digit (Norell & Makovicky, 2004) since this is one of the more noteworthy adaptations of this clade. The appearance of the sickle claw on the second toe in paravians might not be strictly related to a hypercarnivorous and a macrophagous diet, more typical of Eudromaeosauria, but instead to a more generalist and opportunistic behaviour observed in basal Dromaeosauridae (Cau et al., 2015; Cau et al., 2017) and troodontids (Zanno & Makovicky, 2010). This specialisation was then later further exploited in Eudromaeosauria to prey on animals twice as large as the predators (as observed in the “fighting dinosaurs”; Carpenter, 1998), or larger still (Maxwell & Ostrom, 1995).
While the sickle claw was initially considered to have been used to slash or even disembowel prey (Ostrom, 1969), functional studies such as that by Manning et al. (2009) indicate that the manus claws, and by extension perhaps also the second pedal ungual, were better suited for puncturing and gripping prey, or for climbing.
Fowler et al. (2011), based primarily on comparisons between the morphology and proportions of the feet and legs of eudromaeosaurs to several groups of extant birds of prey with known predatory behaviours, suggested that the feet of dromaeosaurids like Deinonychus primarily had a cursorial or grasping function during hunting. In their scenario, large dromaeosaurids could have killed their prey by leaping and subduing them, holding them steady with the help of the sickle claw on digit II, while using the jaws to kill. Forelimbs were covered in long feathers used to flap or keep balance during raptorial behaviour, helped also by the stiff tail acting as a counterbalance: in fact, according to Fowler et al. (2011) claws were capable of exerting considerable force, although the forelimbs had a limited range of motion and more likely were adapted for flapping and not prey manipulation or climbing.
Bishop (2019) conducted a mathematical analysis of the reconstructed 3D models of muscles, tendons, and bones of Deinonychus. The analysis found that the generated claw forces were too weak to slash the soft tissue of prey. On the other hand, coupled with a crouching posture, pedal claws could be effectively used to immobilise or stab the prey. Such a hypothesis is consistent with what is observed in the “fighting dinosaurs” specimen, where Velociraptor is found curled up with its claws penetrating the soft tissues of the Protoceratops in the belly and neck regions.
Bishop (2019) also suggested that the function of the sickle claw could vary among dromaeosaurids. As an example, Kubota & Barsbold (2006) noted that the claw of Adasaurus was relatively smaller when compared with other dromaeosaurids. They therefore suggested that it might have been used with less frequency than in other members of the same clade. One distinct character of the Adasaurus sickle claw is the retained ginglymoid proximal end; investigating the ecology and niche of Adasaurus might give more hints on the function of such peculiar adaptation in more specialised Dromaeosauridae.
Gianechini et al. (2020) highlight a dichotomy among dromaeosaurid dinosaurs, distinguishing those inhabiting Laurasia, specifically velociraptorines, dromaeosaurines, and other eudromaeosaurs, from their counterparts within Gondwana, the unenlagiine dromaeosaurs. This division is manifested most notably in traits related to locomotor and predatory habits. A shorter second phalanx of digit II with a larger heel for the flexor tendon in eudromaeosaurs facilitated the generation of augmented force by the digit. When coupled with a relatively shorter and wider metatarsus it is conceivable that eudromaeosaurs possessed a heightened gripping strength, making them capable of subduing relatively larger prey. Conversely, unenlagiines featured a longer and slenderer metatarsus, likely giving them superior cursorial abilities and enhanced speed. These differences in pedal anatomy may correlate with differences in cranial and dental features. Unenlagiines have long, slender preorbital regions of the skull (Makovicky et al., 2005; Novas et al., 2009) and relatively small, unserrated, and widely spaced teeth compared to eudromaeosaurs, whose more powerfully built crania were capable of generating high bite forces (Gignac et al., 2010). Disparities in the ecological roles and functional adaptations may have contributed significantly to the evolutionary pathways observed in these distinct groups of dromaeosaurs inhabiting the two hemispheres.
A raptorial behaviour and the ability to grapple and hold prey with feet is also supported by the morphology of the hallux. Hattori (2016) noted that in paravians, the articular surface of MT I for MT II has been reduced to a projecting point at the proximal end, similar to that in extant birds. He therefore suggests that MT I could pivot on MT II in paravians. Hattori (2016) reports these anatomical traits were present in Velociraptor. In his interpretation, this adaptation would have allowed the hallux to direct medially, and therefore the forces produced during the flexion of the hallux could be directed perpendicular to that produced by the other digits possibly providing an incipient perching ability and arboreal habits in basal paravians. In derived deinonychosaurians, such morphology could have been co-opted for prey holding, consistently with the previously mentioned hypothesis.
Regarding the rostrum of Velociraptor, Powers et al. (2020, 2021a) found that most Asian and North American eudromaeosaurs could be distinguished by the snout morphology and therefore different ecological strategies: Asian velociraptorinaes have elongated snouts, whereas North American eudromaeosaurs possess a more robust rostrum. The Principal Components Analysis conducted by Powers et al. (2020) on maxillary shape in eudromaeosaurians revealed that Velociraptor possessed relatively long and shallow maxillae, suggesting an adaptation for preying on animals much smaller than itself. This contrasts with taxa like Deinonychus, which had deeper maxillae suited for attacking larger prey. Given this anatomical evidence, it is likely that Velociraptor primarily targeted juvenile dinosaurs, small vertebrates, and opportunistically scavenged larger carcasses.
The overall evidence suggests that Velociraptor was an opportunistic carnivore with a broad dietary range, engaging in both predation and scavenging. While the “fighting dinosaurs” fossil provides a rare glimpse into its active hunting strategy, additional fossil evidence, including bite-marked bones and gut contents (Hone et al., 2010, 2012), underscores the complexity of its feeding behaviour. The presence of velociraptorine teeth associated with various herbivorous remains further reinforces its ecological role as a dynamic predator and a scavenger in the Late Cretaceous ecosystems of Mongolia.
Locomotion
Dromaeosaurids were small to medium-sized obligate bipedal theropods, with an estimated weight ranging between 20 to 80 kg and are typically interpreted as agile lightly-built cursors (Ostrom, 1990; Manning et al., 2009). They are characterised by peculiar anatomical specialisations of the limbs, including: i) highly mobile hand-wrist elements; ii) well-developed raptorial hands with three functional digits; iii) the famous hyperextensible and hypertrophied claw on the second toe of the foot; iv) distinctive caudal vertebrae characterised by highly elongated prezygapophyses, which have been interpreted as structures aiding balance during locomotion and running (Ostrom, 1969, 1990; Manning et al., 2009).
In a recent paper, Gianechini et al. (2020) applied phylogenetic principal component analyses to investigate and quantitatively interpret morphological differences in the hind limbs of eudromaeosaurs and unenlagiines, also in comparison with other theropod groups. According to the authors, the detected morphological differences suggest distinct locomotor and predatory adaptations, with eudromaeosaurs, including Velociraptor, exhibiting features more specialised for grasping and predation rather than sustained cursoriality. Unenlagiines possessed a more elongated and slender metatarsus, a characteristic associated with greater cursorial capacities, allowing for prolonged high-speed locomotion (Gianechini et al., 2020). In contrast, eudromaeosaurs exhibited a shorter, more robust metatarsus, which likely conferred increased grip strength rather than enhanced running abilities (Fowler et al., 2011). The metatarsal structure in unenlagiines suggests adaptations for sustained running, akin to extant falconiforms, whereas eudromaeosaurs, including Velociraptor, had a more compact and mechanically reinforced metatarsus, enabling them to exert a strong grip force (Gianechini et al., 2020), a condition observed in raptorial birds such as owls (Ward et al., 2002; Einoder & Richardson, 2007).
One of the key distinguishing features in eudromaeosaurs is the presence of a robust metatarsus with well-developed ginglymoid distal articular surfaces on metatarsals I, II, and III (Colbert & Russell, 1969; Ostrom, 1969; Norell & Makovicky, 1997, 1999). This morphology suggests a more restricted range of motion in the digits, likely providing increased stability and resistance to torsional stress during prey manipulation (Fowler et al., 2011). In contrast, unenlagiines exhibited a less developed ginglymoid articulation, allowing for a greater range of digital motion, which may have facilitated rapid adjustments during running rather than enhancing grasping strength (Gianechini et al., 2020). These differences highlight a fundamental divergence in general ecological strategies: eudromaeosaurs optimised their hindlimbs for effective prey capture, while unenlagiine’s hindlimbs evolved to enhance locomotor efficiency (Gianechini et al., 2020).
Another significant distinction lies in the proportions of the hindlimb elements. Eudromaeosaurs, such as Velociraptor, displayed relatively shorter tibiae and metatarsi in comparison to the femur, leading to a morphology better suited for sudden bursts of speed rather than endurance running (Gianechini et al., 2020). This configuration aligns with predatory strategies requiring rapid acceleration over short distances, possibly for ambush hunting or close-range attacks. By contrast, unenlagiines possessed a longer tibia and an elongated subarctometatarsalian metatarsus, features indicative of improved stride efficiency and lower energetic cost during sustained locomotion (Carrano, 1999; Fowler et al., 2011; Gianechini et al., 2020). These differences suggest that while unenlagiines may have been capable of higher overall velocities and prolonged chases, eudromaeosaurs were likely adapted for short, powerful sprints aimed at subduing prey (Gianechini et al., 2020).
Additionally, the pedal phalanges of eudromaeosaurs reflect their predatory adaptations (Gianechini et al., 2020). The elongation of the distal pre-ungual phalanges in Velociraptor and its relatives is comparable to extant raptorial birds with strong grasping abilities, such as Bubo and Turdus (Fowler et al., 2011). This trait, along with a shorter phalanx II-1 and more pronounced interphalangeal ginglymoid articulations, suggests that eudromaeosaurs relied on their feet for prey restraint, possibly using their second pedal digit as a lethal gripping tool (Ostrom, 1969; Fowler et al., 2011). Unenlagiines, in contrast, exhibited more gracile phalanges with reduced ginglymoid articulation, further supporting the hypothesis of a cursorial rather than grasping-oriented function (Gianechini et al., 2020).
A comparative analysis with extant raptorial birds supports the inference that eudromaeosaurs, like Velociraptor, prioritised grasping strength over cursorial efficiency. Birds with robust and shorter metatarsi, such as owls, exhibit powerful grip forces, whereas those with elongated metatarsi, like falcons, emphasise speed and agility (Ward et al., 2002; Einoder & Richardson, 2007; Fowler et al., 2009). Similarly, Velociraptor’s hindlimb proportions indicate a biomechanical advantage in forceful prey restraint rather than endurance running. These adaptations highlight a trade-off between cursoriality and grasping efficiency, with unenlagiines evolving towards greater terrestrial mobility and eudromaeosaurs refining their hindlimbs for specialised predation (Gianechini et al., 2020).
Gianechini et al. (2020) concluded that the morphological differences between the hindlimbs of unenlagiines and eudromaeosaurs, including Velociraptor, underscore their distinct ecological roles. Unenlagiines likely excelled in cursorial locomotion, with an elongated and slender metatarsus allowing for sustained high-speed movement. In contrast, eudromaeosaurs exhibited a more compact, reinforced hindlimb structure optimised for grasping and prey restraint, at the expense of prolonged running efficiency.
The tail of Velociraptor was a key anatomical feature that played a crucial role in their locomotion and balance (e.g., Ostrom, 1969; Holtz, 2003; Norell & Makovicky, 2004) (Fig. 6). Nonetheless, it was not entirely inflexible allowing some important minor movements, and some complete specimens are preserved with curving tails that retain their ossified tendon bundles in articulation (e.g., MPD-C 100/986). The primary function of a rigid tail in Velociraptor was likely to maintain balance while running at high speeds. In a fast and agile predator, a well-balanced tail would have helped make sharp turns while running. The tail also may have contributed to stability during various activities, such as stalking, pouncing, and grappling with prey. When engaged in predatory behaviour, maintaining balance was probably essential to avoid injury and ensure a successful hunt. Moreover, a horizontally held, stiff tail might have also reduced air resistance while running. This streamlined posture could have made Velociraptor a more efficient runner, conserving energy during fast moving. Increased agility, perhaps in part due to this peculiar tail, in Velociraptor mongoliensis is indicated by the relatively large size of the floccular lobes in the endocranium (King et al., 2020).
Metabolism
Tada et al. (2023) recently used Velociraptor mongoliensis (MPC-D 100/2000) nasal cavity and brain volumes as proxies to assess its degree of endothermy/ectothermy. The authors found different relationships between nasal cavity size and skull/head size in endothermic taxa versus ectothermic ones in a sample of living amniote taxa. Their analysis found Velociraptor mongoliensis to plot between the endothermic and ectothermic best-fit lines. However, without a measure of uncertainty in the form of confidence intervals or similar metrics, it is unclear whether this intermediate position is significant or not. Although this method is based on a single proxy, it is consistent with the results of some other studies: Legendre et al. (2016) estimated resting metabolic rates in a range of living and extinct archosaurs from the sizes and densities of osteocyte lacunae, and found values above that of extant ectotherms, but below those for extant birds, for Troodon formosus, a close paravian relative of Velociraptor. If nothing else, the results from Tada et al. (2023) indicate that Velociraptor exhibits at least one trait consistent with reptilian thermophysiology, specifically a relatively small nasal cavity although, as Legendre et al. (2016) noted, the division of taxa into discrete ectothermic and endothermic bins based on the physiology of living species may be too crude to capture the variation and its significance in fossil forms.
Comparing encephalisation quotients, although this is not a widely accepted proxy for metabolic rates, coelurosaurs were suggested to be as active as many birds and mammals, whereas less derived theropods and ornithopods were slightly less active than birds and mammals, but more active than extant reptiles (Hopson, 1980; Fastovsky & Weishampel, 2009). Conversely to the results of Tada et al. (2023), Seymour et al. (2012) found that foramen size and blood flow in extant mammals are directly proportional to metabolic rates, and therefore reported similar dinosaur large foramina should indicate a routinely highly active and aerobic lifestyle, possibly linked to endothermy. One more aspect regarding Velociraptor is that, as in other maniraptorans, some ribs possessed an uncinate processes on the posterior margin. These are proportionately as long as in extant diving birds and their length was found to be related to bird’s oxygen requirements (shorter in walking birds, longer in diving birds); this would suggest that some maniraptorans had high oxygen requirements (Tickle et al., 2007; Codd et al., 2008; see also Fisher et al., 2000). Also, the presence of air sacs in theropods (e.g., Naish et al., 2004; O’Connor & Claessens, 2005) can be interpreted as evidence of an increased respiratory capacity and efficiency, linked to a high metabolic rate. Air sacs might have played a role in thermoregulating body temperatures as well. Indeed, the presence of feathering itself, possibly functioning as an insulator, can be interpreted as further evidence for endothermy, or at least as evidence of elevated metabolic rates.
Consequently, even though, Tada et al. (2023) results tend to show an intermediate condition between the reptilian and the avialan physiologic condition, other evidence (of which here we provide only some spare examples) tends to suggest that the condition of dinosaurs, especially derived theropods, was closer to extant bird’s endothermy. Yet we have to consider that the avialan endothermic state (which varies even between living species) must have been reached through a series of evolutionary steps along a continuum. Numerous proxies support an elevated metabolic rate relative to extant reptiles, but it is not clear if Velociraptor had rates comparable to extant birds. A “mosaic” of adaptations, many of them more similar to the avialan endothermic state and few to the reptilian ectothermic state (such as a small nasal cavity), is expected.
Social behaviour: pack hunting and living in groups
Several lines of evidence indicate that dromaeosaurids were gregarious dinosaurs. Maxwell & Ostrom (1995) interpreted the taphonomy of several Tenontosaurus-Deinonychus associations, including the Deinonychus type locality as showing evidence of multiple specimens of Deinonychus hunting and feeding on the much larger ornithopod taxon. Li et al. (2007) reported a series of trackways from Shandong, China, where the trackmakers were suggested to be large-sized eudromaeosaurs. The parallel trackways were made by six individuals of the same ontogenetic stage, walking at a slow pace and likely moving together in the same direction. According to the authors, this would suggest that at least some species of dromaeosaur lived in groups, albeit the trackways do not provide evidence for pack hunting. This observed habit offers a glimpse into the potential gregarious tendencies of dromaeosaurs, prompting questions about the factors and benefits that travelling in groups may have conferred. The coexistence of dromaeosaurs within the same ontogenetic stage in group settings raises questions also about their ecological dynamics. Such group living arrangements might imply instances in which individuals of similar ontogenetic stages might have opportunistically targeted the same prey. This does not necessarily imply well-coordinated hunting strategies or habitual pack hunting, but rather the possibility of chance encounters leading to the convergence of hunting interests. This is consistent with the finding of associated skeletons of Deinonychus and one Tenontosaurus (Maxwell & Ostrom, 1995; Ostrom, 1969, 1990), supporting a case of opportunistic feeding behaviour (Brinkman et al., 1998), or solitary hunters that occasionally foraged in loose, ephemeral associations (Roach & Brinkmann, 2007).
A study by Frederickson et al. (2020) analysing oxygen isotopes in teeth demonstrated that Deinonychus shows ontogenetic diet partitioning, revealing therefore a distinct behaviour from cooperative social mammals. This investigation challenges the preconceived idea of pack hunting and uniform dietary strategies in dromaeosaurs. The authors also found that juvenile and adult teeth were often associated together and hypothesised that, due to their lack of spatial separation from adults, these dromaeosaurs likely employed hunting techniques comparable to those of many avian raptors (Ellis et al., 1993). This would also exclude a Komodo dragon-like behaviour (as speculated by Roach & Brinkmann, 2007) where high levels of intraspecific aggression and cannibalism can be observed and where juveniles actively avoid adult predators (Foster et al., 1988; Werner et al., 1983; Keren-Rotem et al., 2006). On the other hand, Frederickson et al. (2020) suggested that it is possible that dromaeosaurs showed a more ratite-like parental care relationship.
Speaking strictly of Velociraptor, there is no particular evidence to state that this taxon generally behaved differently from Deinonychus, as depicted in the previous paragraphs, although different size and snout morphologies between the two suggest different prey acquisition methods (Powers et al., 2021a). According to Powers et al. (2021a), a robust snout (such as in Deinonycus) would have been beneficial in handling larger prey items. On the other hand, velociraptorines show snout morphologies more like modern small prey specialists, consistent with their ecosystem (Jerzykiewicz et al., 2021). Nonetheless, they were not limited to small vertebrate prey, as exemplified by the “fighting dinosaurs” specimen (Powers et al., 2021a). Other examples of scavenging and or predatory behaviours in Velociraptor were given by Hone et al. (2012), where bones of an azhdarchid pterosaur were found in the stomach content of MPC-D 100/54, and teeth referred to Velociraptor were found with protoceratopsid bones (Hone et al., 2012). The latter find is from the Bayan Mandahu locality, where ‘Velociraptor’ osmolskae was collected. This nuanced understanding of snout morphology, dromaeosaurs’ habits, and prey selection underscores the versatility of this dinosaur group in preying on very different animals, generally actively hunting small and scavenging on larger prey.
Palaeaoneurology
The only palaeoneurological study applied to Velociraptor derives from the analysis of a digital endocast of specimen MDP 100/976 by King et al. (2020). The specimen was collected during the 1991 expedition of the Mongolian Academy of Sciences and the American Museum of Natural History from the Djadokhta Formation at Tugrugeen Shireh, Mongolia (Norell et al., 1997). MPC-D 100/976 consists of a partial skeleton including an incomplete neurocranium comprising the prootics, exoccipitalsopisthotics, supraoccipital, basioccipital, and basisphenoid (Norell et al., 2006; King et al., 2020). It was identified as Velociraptor mongoliensis (e.g., Norell et al., 1995, 2006; King et al., 2020).
The virtual brain endocast of MPC-D 100/976, obtained from high-resolution tomographic scan, revealed areas of the medulla oblongata, cerebellum, cerebellar flocculi, and the entire basiencephalic surface corresponding to these elements, including the labyrinth of the inner ear on both sides. Among the most significant elements in the brain of MPC-D 100/976 are the cerebellar flocculi and their relatively large size compared to the rest of the endocast. The flocculi, or floccular lobes, are involved in maintaining the stability of the head and eyes during movement, indicating the agility of a vertebrate (Witmer & Ridgely, 2009). The large size of the floccular fossae, in the case of dromaeosaurids like Velociraptor and more generally maniraptorans, can be associated with a pronounced sensitivity in balance regulation during locomotion (King et al., 2020), as observed in modern non-flying birds, where the hypertrophy of the floccular lobes appears to be related to stabilising the inherently unstable nature of bipedal locomotion (Walsh et al., 2013). The hypertrophy of cortical regions such as the cerebellar flocculi explains how fast movements and gaze stabilisation were essential to the life habits of Velociraptor (King et al., 2020). Large cerebellar flocculi are also correlated with enhanced vestibulo-ocular (VOR) and vestibulocollic (VCR) reflexes (Hopson, 1977; Witmer & Ridgely, 2009), suggesting that Velociraptor was capable of easily tracking moving objects (King et al., 2020). This thesis seems to be reinforced by osteological data on orbital dimensions and semicircular canals in the species (Stevens, 2006; Schmitz & Motani, 2011; Torres & Clarke, 2018).
Another element of interest is the architecture of the inner ear. In life, the endosseous cochlear duct of Velociraptor would have housed the basilar papilla, the auditory organ of tetrapods (Gleich et al., 2005; Walsh et al., 2009), where the relationship between duct length and papilla size allows this bony element to be used to indirectly estimate the ability to perceive medium and high-frequency sounds in non-avian dinosaurs (Witmer & Ridgely, 2009; Lautenschalager et al., 2012). It has also been demonstrated that the relationship between the length of the cochlear duct and the basisphenoid is correlated with auditory frequencies in modern archosaurs (Walsh et al., 2009), providing a method for estimating sensitivity to medium and high frequencies in non-avian dinosaurs. Data on the dimensions of the cochlear duct and basisphenoid in MPC-D 100/976 indicate a frequency range centred around 2,368 Hz with an upper limit for high frequencies of 3,965 Hz (King et al., 2020). This is comparable to the range of hearing in modern birds such as the raven (Corvus corax) and the African penguin (Spheniscus demersus) (Walsh et al., 2009), suggesting an auditory perception range more similar to that of current neognaths than that of basal archosaurs and other reptiles. Velociraptor likely had the ability to hunt, hear, and perhaps vocalise efficiently in the range of 2,400 Hz (King et al., 2020).
The elongated structure of the cochlear duct in Velociraptor indicates that this species was capable of perceiving a wide range of sounds, indicating that hearing was a sense of significant importance in this species (Manley, 1990; Walsh et al., 2009; Witmer & Ridgely, 2009; Brusatte et al., 2016; Carabajal et al., 2016). King et al. (2020) also proposed that Velociraptor could have used hearing in social interactions as well as active predation, since the size of the cochlear duct of MPC-D 100/976 falls within the range shown by Melopsittacus undulatus (budgerigar), a socially vocal learning species. The neuroanatomical evidence of MPC-D 100/976, particularly the link between motor performance and floccular cerebellar size, is supported by the observations of Witmer & Ridgely (2009).
Although Choiniere et al. (2021) highlight the prominent auditory and visual performance of theropods from morphological and functional observations on cranial elements, accompanied by the relative volumes of certain neurocranial regions, few direct parallels can be drawn for Velociraptor using the partial brain of MPC-D 100/976. However, the detailed evidence from the specimen’s auditory canal, along with the vestibulo-ocular and vestibulocollic performance deduced from the neuroanatomy, do not question the refined visual and auditory sensitivities in Velociraptor. Only a detailed analysis of a more complete endocast can unequivocally clarify its sensory potential, additionally supporting or refuting the possible nocturnal habits hypothesised by Schmitz & Motani (2011) (see also Motani & Schmitz, 2011) based on orbit dimensions.
Palaeopathologies
The study of palaeopathologies, their causes, nature, and healing can provide insights into the autecology of the animals that suffered the injury (e.g., Bertozzo et al., 2017, 2021). However, there are relatively few reports of palaeopathologies in Velociraptor other velociraptorines.
MPC-D 100/54, a young individual referred to Velociraptor mongoliensis, was reported to have a broken and partially healed rib showing signs of regrowth (Hone et al., 2012). The Velociraptor mongoliensis MPC-D 100/976 braincase shows a reported unhealed bitemark in the form of punctures on the frontal bone. It was considered to be caused by an intraspecific agonistic interaction since the punctures show the spacing between neighbouring teeth of a specimen roughly the same size as MPC-D 100/976 (Norell et al., 1995; Norell & Makovicky, 1999; Molnar, 2001). The unhealed nature of this bitemark strongly suggests that it proved fatal, shedding light on intraspecific interactions and potential confrontations within the species.
Among other velociraptorines, the holotype of Adasaurus was initially suggested to represent a largely pathologic individual, possibly due to serious injury or a specific disease (Norell & Makovicky, 1997, 2004). However, a re-examination of the specimen by Turner et al. (2012) determined that the holotype was non-pathological. Instead, certain periosteal features were attributed to advanced age rather than injuries. This revaluation highlights the significance of careful research in separating bone pathologies from age-related changes in dinosaur remains.
Roach & Brinkman (2007) published photographs of a pathological phalanx II-2 of Deinonychus, which healed with a large offset between proximal and distal sections following a fracture. They interpreted this pathology as being consistent with pedal digit II encountering significant stresses when subduing prey, leading to injury in one individual.
Recent findings by Jasinski et al. (2022) regarding Dineobellator have unveiled evidence of re-growth and remodelling in several skeletal elements, including ribs, premaxillary fragments, caudal vertebrae, and manual unguals. In particular, one of the ribs shows an abnormal bone growth in its proximal region.
In summary, while the palaeopathological evidence reported in velociraptorines does not give a complete picture of their behaviour, it does provide some clues about their resilience to injuries, intraspecific interactions, changes with age, and possibly on the use of the hallmark second pedal claw.
Palaeoenvironments
The palaeoenvironment of Velociraptor can be reconstructed based on the sedimentological features of the Djadokhta Formation at the Flaming Cliffs locality in Mongolia. Comparisons between the Djadokhta and the overlying Nemegt Formation highlight significant differences in palaeoenvironmental conditions, with the first reflecting a harsher, drier setting, while the second one indicating a more humid, fluvial-dominated environment capable of supporting dinosaur species of larger size (Clarke & Norell, 2004). In particular, the fossiliferous deposits of the Djadokhta Formation primarily consist of poorly cemented, fine-grained arkosic sandstone with large-scale cross-stratification and occasional caliche horizons, indicating aeolian dune deposits and associated playa lake sediments (Gradziński et al., 1977; Ostrom, 1990). These sedimentary features suggest that the region experienced at least semi-arid conditions, indicating possible high plateau landscapes. The palaeoenvironment was likely shaped by episodic climatic variability, where arid periods dominated, interspersed with occasional wetter intervals that facilitated the formation of ephemeral water bodies (Dashzeveg et al., 2005; Novacek et al., 2022). These transient aquatic environments may have played a crucial role in sustaining the local fauna, including Velociraptor. As a whole, the low diversity of vertebrate assemblages, combined with the small body sizes of many dinosaur species, implies an ecosystem in which resource competition was significant, potentially influencing the predatory behaviour and ecological adaptations of theropods (Godefroit et al., 2008).
Additionally, the sedimentological evidence from the Flaming Cliffs suggests that sandstorms were a common occurrence, as indicated by the presence of well-preserved and articulated dinosaur skeletons buried in structureless sandstones (e.g., the already mentioned famous “fighting dinosaurs”). These specimens, often found in lifelike poses, imply that some individuals perished while attempting to free themselves from sandstorm deposits (Jerzykiewicz et al., 1993).
The taphonomic evidence from Velociraptor fossils may provide further evidence to reconstruct its peculiar palaeoenvironment of life. In particular, fossilised bones exhibit trace marks attributed to scavenging activity by insects and small mammals, particularly on limb joints where collagen-rich cartilage was present (Saneyoshi et al., 2011). The high frequency of these traces suggests that in the nitrogen-poor desert environment of the Late Cretaceous Gobi Desert, dried dinosaur carcasses served as a critical nutritional resource for smaller organisms (Saneyoshi et al., 2011). Additionally, the rapid burial of some dinosaur remains within aeolian deposits suggests that sandstorms played a significant role in fossil preservation, allowing for exceptional skeletal articulation and the preservation of delicate anatomical features (Fastovsky et al., 1997).
The broader palaeoenvironmental context of the Djadokhta Formation can be understood within a regional framework, with depositional environments at sites such as Tugrikin Shireh and Bayan Mandahu exhibiting strong similarities; this indicates that the southern Gobi Desert during the Late Cretaceous consisted of geographically limited basins formed by block faulting (Fastovsky et al., 1997). These basins likely contained a combination of aeolian dune fields and alluvial runoff zones, suggesting a complex mosaic of depositional environments that shaped local ecosystems (Eberth, 1993). Such a setting would have necessitated behavioural adaptations in Velociraptor, potentially including a high degree of territoriality and an opportunistic feeding strategy to cope with fluctuations in prey availability.
Thus, considering all the sedimentary and taphonomic evidence, the palaeoenvironment of Velociraptor, as inferred from the Djadokhta Formation, reveals a complex and challenging habitat marked by arid to semi-arid conditions, episodic moisture availability, and frequent sandstorm events. It follows that Velociraptor thrived in a resource-scarce ecosystem where adaptability, efficient predation, and scavenging behaviours represented very likely key survival strategies.
VELOCIRAPTOR ICONOGRAPHY: EVOLUTION OVER TIME AND INFLUENCE ON POPULAR CULTURE
Over the years, Velociraptor has been reconstructed by numerous palaeoartists, with varying degrees of accuracy (Manucci & Romano, 2023) (Fig. 8). Despite its status as one of the most depicted and popular dinosaurs, it remained largely unknown to the general public for over half a century, until the discovery of the “fighting dinosaurs” in 1971. This exceptional specimen is a true fossil treasure for both science communication and paleoart, as it captures a moment in time, providing an ideal basis for inferring interactions between Mesozoic species and for artistic reconstruction of those interactions. As Barsbold (1998) stated, “The Fighting Dinosaurs are among the most unique examples in the paleontological record ever discovered during the long history of this science”.
- Iconographic history of Velociraptor through the last 50 years.
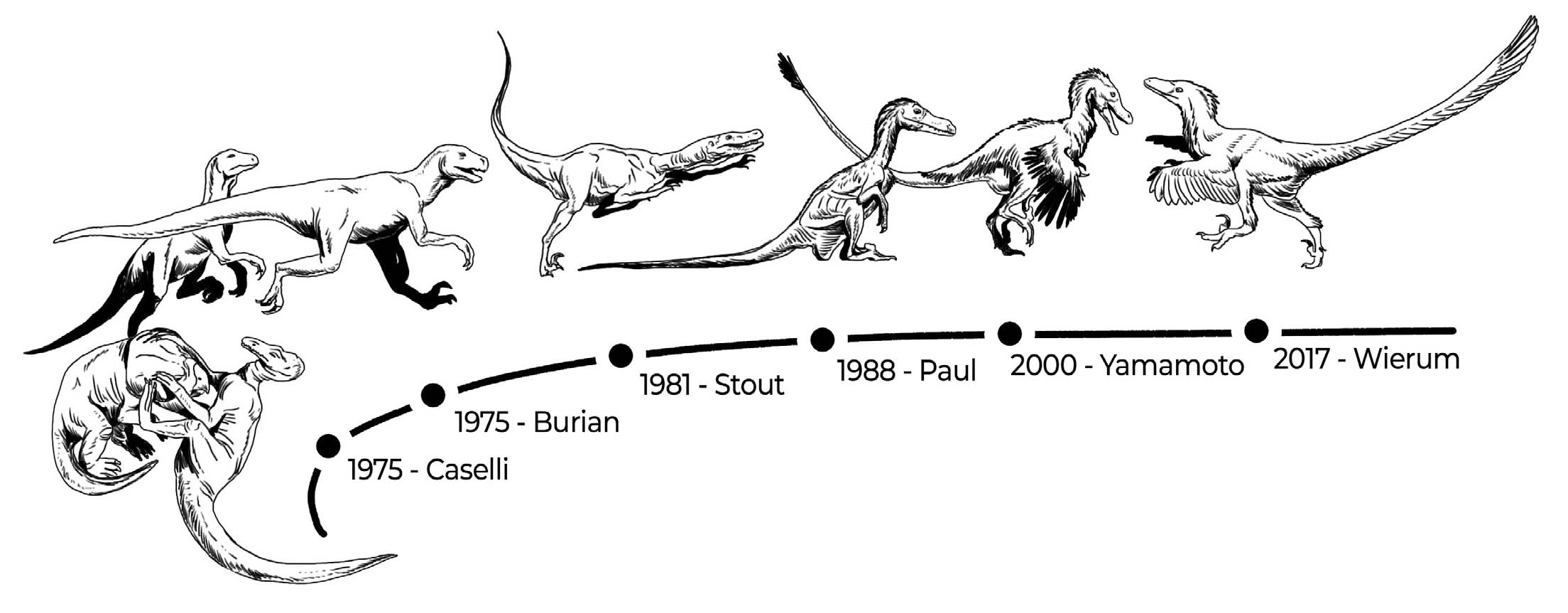
The first reconstruction of Velociraptor, illustrating its battle with Protoceratops, appeared in 1975 in a book illustrated by Giovanni Caselli (Halstead, 1975), which featured many new discoveries and previously overlooked taxa, including the first ‘invivo’ reconstruction of Spinosaurus. However, Caselli’s Velociraptor lacked the characteristic sickle claw on its foot. In the image, Protoceratops passively endures the attack, and Halstead (1975) emphasises the prey’s vulnerability in the text, noting that “Its beak was adapted for cutting tough plants, not for fighting”. This interpretation is contradicted by the fossil itself, in which the right forearm of the Velociraptor is grasped by the clenched beak of the Protoceratops, effectively preventing the predator’s escape.
The same year, the well-known palaeoartist Zdenek Burian also illustrated Velociraptor, portraying it in a dynamic pose, but reconstructing its head with proportions more consistent with Deinonychus, a mistake repeated by the Jurassic Park franchise. This error was partly due to the difficulty in obtaining reference materials, as the available photographs showed a partially prepared stage of the fossil, and the specimen still lacks a comprehensive study detailing all its skeletal elements, despite being one of the most famous fossils in the world. Burian likely relied on the renowned monograph on Deinonychus (Ostrom, 1969), which was richly illustrated, and he assumed the two animals were similar, even though the outline of the Velociraptor skull was well-known since the paper by Osborn (1924). It is noteworthy that Burian chose to depict the animal alongside another reptile from the Djadokhta Formation, the squamate Gilmoreteius, a genus for which an osteological study had just been published in 1975 (Sulimski, 1975).
Other reconstructions from this period include works by artists such as Gregory Irons (Long, 1978) and William Stout, both of whom were also active in projects related to comics and animation. This period marks the entry of dromaeosaurids into the collective imagination, primarily due to the book illustrated by Caselli (Halstead, 1975), which served as an inspiration for the comic “Flesh” (1977-2016) by Pat Mills (Liston, 2010). The comic featured Deinonychus, portrayed dynamically and inspired by the latest discoveries. William Stout (Stout, 1981) would later inspire many subsequent pop creations; one of these, “The Natural History Project” by Jim Henson of 1986), was intended to be an animated film inspired by Stout’s palaeoart, but the project was never completed and was eventually abandoned. Among Stout’s concept art was Velociraptor, in a version that even included speculative sparse plumage, albeit in a caricatured and grotesque style.
During the 1980s, more accurate reconstructions emerged, thanks to the rigorous work of palaeoartists such as Mark Hallett (Waldrop & Loomis, 1984) and Gregory Paul (Paul, 1988), who were also the first to envision Velociraptor as fully feathered, anticipating the discoveries of feathered dromaeosaurids from China in the following decades. Paul proposed the first skeletal drawing, and his volume “Predatory Dinosaurs of the World” (Paul, 1988) soon became the reference that transformed Velociraptor into a popular star. This anatomical diagram, presented in multiple perspective views and inspired by the fighting dinosaur specimen, remains one of the best visual references for reconstructing the animal and was immediately adopted as a model by many other palaeoartists. Other notable but more conservative reconstructions from this period were realised by Renzo Zanetti (Bonaparte et al., 1984), Douglas Henderson (Wilford, 1985), John Sibbick (Norman, 1985; Gardom & Milner, 1993), and Steve Kirk (Dixon et al., 1988), where the skin and limb musculature were more inspired by those of living reptiles. In some “intermediate” versions, limited coverage of proto-feathers along the back or head was imagined, as in the reconstructions by Wayne Barlowe (Preiss & Silverberg, 1992) and Brian Franczak (Wallace, 1993). Some new reconstructions inspired by the fighting dinosaurs, particularly those by Hallett (Czerkas & Czerkas, 1990), Sibbick (Gardom & Milner, 1993), Luis Rey (Rey, 2001), and Raul Martin (Chiappe, 2003; Fig. 9) proposed a scenario where the predator had to defend itself from Protoceratops, which was no longer as harmless as initially imagined by Halstead (1975), a view also consolidated in a study on the specimen by Carpenter (1998).
- The “fighting dinosaurus”, artwork by Raul Martin, from Chiappe (2003).
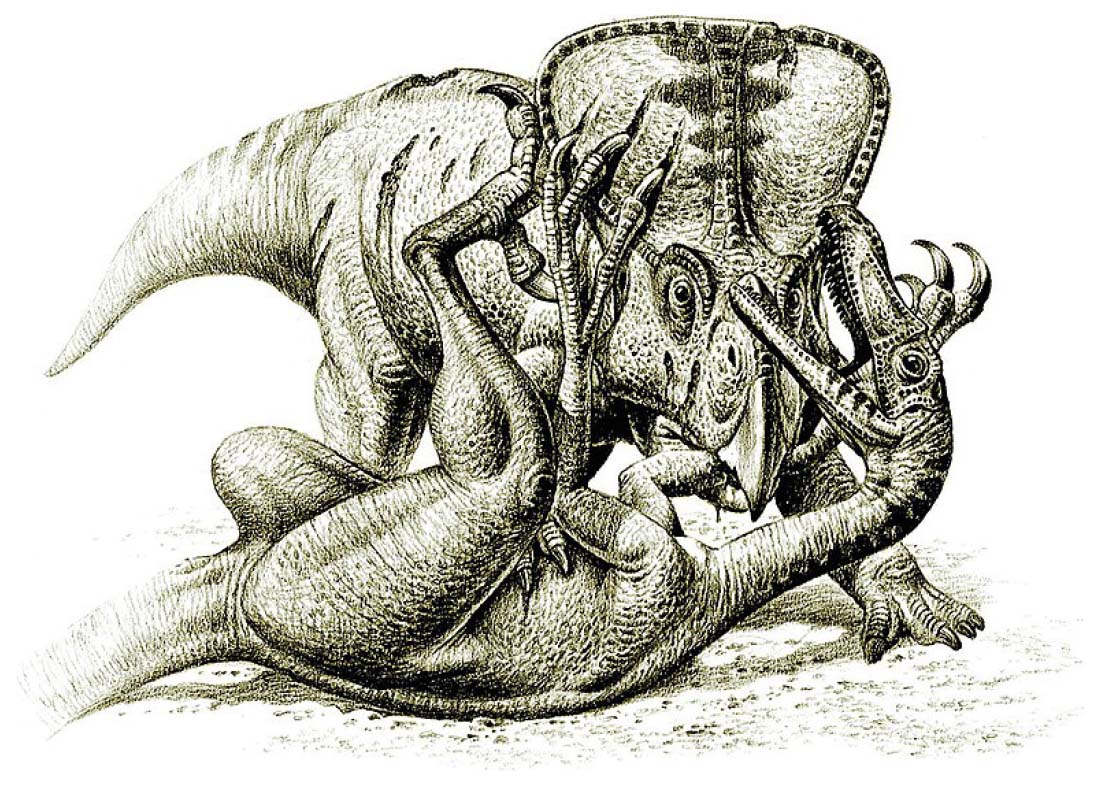
Many of these artists were featured in the exhibition “Dinosaurs Past and Present” (1986), curated by Stephen and Sylvia Czerkas, and among the visitors to the inaugural edition in Los Angeles was the writer Michael Crichton, who discovered the potential of the new dinosaur discoveries through this event. The first popular debut for Velociraptor is due to the well-known novel “Jurassic Park” by Michael Crichton published in 1990, and especially to its film adaptation by director Steven Spielberg, released in 1993. In both the book and the impactful film, these dinosaurs are portrayed as fearsome, intelligent predators, hunting in packs. The “raptors” in the movie are about twice the actual size and are inspired by Deinonychus, as Gregory Paul himself declared in an article recounting his role as a scientific advisor for the film (Paul, 2017), for which he realised various anatomical diagrams. The erroneous use of the name is based on the taxonomy proposed by Paul (1988), which was one of the references for the novel and in which Velociraptor was suggested as the sole genus for both Velociraptor mongoliensis and Deinonychus antirrhopus. Despite Paul (1988) and others (e.g., Bakker, 1986) hypothesising extensive feathering in dromaeosaurids, based on phylogenetic grounds, there was no supporting fossil evidence for it until the discovery of Sinornithosaurus (Xu et al., 1999). At the time of the film’s release, there was, in fact, no direct evidence of the presence of feathers in these theropods. In 2007, small marks that resemble the attachment sites of wing feathers in birds were identified on the ulna of a Velociraptor specimen, thus indicating that this dinosaur likely had feathers (Turner et al., 2007).
These groundbreaking discoveries have not yet been fully embraced by popular culture, which is still dominated by cinematic portrayal, as a quick Google Images search can demonstrate. Although Velociraptor has gone from being obscure to one of the most recognisable dinosaurs to the public, it has also remained one of the most misunderstood. For over 20 years, no palaeoartist has depicted this animal with partial feathering, in line with fossil evidence. However, before these discoveries, this vision ofthe animal attracted scepticism even among researchers, as palaeoartist Luis Rey, one of the first to promote the new image of feathered dromaeosaurids, wrote in 1996: “I have suffered somewhat with my restorations of feathered dinosaurs, becoming blacklisted by the very paleontological community here in England. However, every time they want a ‘fantasy’ dinosaur, they remember me. Sad, really, because I try to be as rigorous as possible in my anatomical reconstructions and I tend to honor requests to draw featherless theropods” (Debus & Debus, 2002). During the 1990s, Luis Rey was also among the first to sculpt a life-sized model of Velociraptor with feathers (Rey, 2001); the subsequent discovery of Sinornithosaurus would confirm how these early, “extreme” reconstructions (without wings) were actually cautious compared to what was later revealed by the fossils themselves. In some cases, even the popular image has undergone significant updates. In the Disney movie “Dinosaur” (2000), the inspiration returned to the actual genus Velociraptor, instead of Deinonychus, and various palaeoartists were involved in the realisation of the preparatory studies. David Krentz proposed a design for Velociraptor that included feathers, unfortunately discarded due to the greater complexity of 3D rendering. Advances in technology and the discovery of an increasing number of feathered species have led to many innovations: the documentary series “Dinosaur Planet” (2003), supervised by Scott D. Sampson, dedicates an entire episode to Velociraptor and its ecosystem, finally reconstructing the animal with dense plumage. In 2011, it was depicted in the series “Dinosaur Revolution,” again led by palaeoartist David Krentz. Recently, an even higher level of realism was achieved in the series “Prehistoric Planet” (2022). Once again, this quality was made possible by involving palaeoartists in the creative team, including Gabriel Ugueto, Mark Witton, and David Krentz, along with Scott Hartman for skeletal reconstructions (which include Velociraptor) and the palaeontologists Darren Naish, John Hutchinson and Steve Brusatte; in many other documentary productions, this does not occur, sometimes resulting in representations more inspired by cinema than palaeontology. Behaviourally, all these series depict Velociraptor as gregarious and hunting in small groups. Partly this derives from initial hypotheses about social hunting in Deinonychus (Ostrom, 1969), the plausibility of which has subsequently been questioned by taphonomic evidence and is now of dubious value (Roach & Brinkman, 2007). The hypothesis that they temporarily formed small groups is not impossible, especially considering the varied social behaviours among closely related animal species today, and the hypothesis has been suggested again to explain some track sites where multiple individuals likely moved together (Li et al., 2007), as supported in other dinosaur groups. Citing the study by Li et al. (2007): “Because the sedimentology indicates a relatively rapid emplacement and subsequent burial of the trackways, a scenario in which six similarly sized individuals independently generated parallel, regularly spaced, non-overlapping trackways is highly unlikely”. Only further discoveries will provide a more complete idea of the ethology of different dromaeosaurids, confirming or disproving the hypothesis of a solitary predator or, conversely, a broader spectrum of behaviours.
A hundred years after its discovery, new fossils and analyses have revealed the most probable appearance of Velociraptor. Although direct evidence of feathers and pack-hunting remains elusive, it is doubtless that future discoveries and new methods will continue to reveal new aspects of Velociraptor biology as palaeontologists labour to bring this ancient predator “back to life”.
Reconstruction of Velociraptor
Figure 10 shows our updated reconstruction of Velociraptor mongoliensis. The plumage on the neck is inspired by that of Sinornithosaurus, Daurlong (Wang et al., 2022), and other paravians (Li et al., 2010; Lefèvre et al., 2017; Hu et al., 2018; Poust et al., 2020), which in various parts of the body are covered by down or ratite-like feathers, a much simpler plumage compared to that found on the wings of extant birds. In some genera, the neck plumage consists of both pennaceous and down-like feathers, as portrayed in our reconstruction. We chose a colouration with brownish-red tones, which is common in many terrestrial birds and in fossilised coelurosaurs, where preserved melanosomes and melanin were fossilised (Zhang et al., 2010; Li et al., 2014). Our choices are consistent with the inferred exclusively terrestrial habits, just like in ratites, and in contrast to some dromaeosaurids that were more adapted for glide and whose plumage is also known (Li et al., 2012). The presence of scales on the snout currently lacks direct evidence, as does the presence of a covering over the teeth, based on a series of clues and comparisons with modern reptiles (Cullen et al., 2023). The crests in front of the orbits, often depicted in past reconstructions (Paul, 1988), were excluded from our reconstruction, being incorporated into a continuous tissue (supraorbital membrane) from the lacrimal to the postorbital (Maidment & Porro, 2009; Snively et al., 2013), as typically observed in modern sauropsids.
- Photogrammetric scan of a Velociraptor cast (MUST SN1140/BM) and digital reconstruction of the hypothesised in-vivo appearance of Velociraptor mongoliensis.

CONCLUDING REMARKS
The centenary of the first description of Velociraptor offers a unique opportunity to reflect on the extensive journey of research, interpretation, and cultural integration surrounding this iconic theropod dinosaur. Over the past century, Velociraptor has transformed from a relatively obscure fossil to a household name, largely due to its prominent role in popular media. However, its journey through scientific understanding has been equally transformative, revealing complex insights into its biology, behaviour, and ecology.
The first Velociraptor fossil, discovered on August 11, 1923, by a team led by Roy Chapman Andrews from the American Museum of Natural History (AMNH) in the Gobi Desert, marked the beginning of a century of palaeontological exploration and discovery (Gallencamp, 2001). The crushed skull and manual claw found in the Djadokhta Formation at the Flaming Cliffs were pivotal in defining Velociraptor distinctive anatomy and predatory capabilities.
Initially, artistic portrayals of Velociraptor were somewhat limited, with early reconstructions failing to fully capture its unique features, such as the sickle claw. The discovery of nearly complete skeletons and iconic fossils like the famous “fighting dinosaurs” specimen in 1971, which depicts a Velociraptor in combat with a Protoceratops, significantly advanced our understanding of its anatomy and behaviour. This specimen highlighted possible predatory tactics and social interactions in Velociraptor, challenging earlier assumptions about its behaviour.
The late 20th and early 21st centuries brought a paradigm shift in our understanding of Velociraptor and dromaeosaurids in general. The discovery of feathered dromaeosaurids, such as Sinornithosaurus, and the subsequent hypothesis that Velociraptor also had feathers, drastically changed its depiction from a scaly predator to a feathered, bird-like dinosaur. This shift was further supported by evidence of feathers in closely related species and anatomical features in Velociraptor itself, such as the presence of quill knobs on the forearm, suggesting attachment points for feathers.
In last decades, modern technologies have greatly enhanced our understanding of theropod dinosaur anatomy and biology. Advanced imaging techniques have allowed palaeontologists to study the internal structures of Velociraptor fossils, leading to new insights into its physiology and sensory capabilities. For instance, studies of its braincase have revealed an enlarged olfactory bulb, indicating a keen sense of smell, whereas analyses of its inner ear suggest advanced auditory capabilities. These findings suggest that Velociraptor very likely was a highly perceptive predator, capable of sophisticated hunting strategies.
The distinctive sickle claw of the taxon has been a focal point in understanding its predatory behaviour. The prevailing hypothesis suggests that Velociraptor used their feet for grasping and immobilising prey rather than slashing, a theory supported by the biomechanics of its limbs and the positioning of its claw. This is exemplified by the “fighting dinosaurs” fossil, where the Velociraptor pedal claw is seen in a gripping/kicking motion against its prey. Such morphological adaptations highlight its specialisation in capturing and subduing prey, reflecting its ecological role as an efficient predator.
Feeding strategies in Velociraptor likely included a mix of active hunting and scavenging, as suggested by stomach contents and evidence of varied prey. This versatility in feeding strategies indicates a flexible approach to survival in the harsh environments of Late Cretaceous Mongolia. Additionally, while there is limited direct evidence of pack hunting, trackway studies and comparisons with other dromaeosaurids suggest that Velociraptor might have exhibited some form of social behaviour, possibly for cooperative hunting or social interactions.
Velociraptor’s depiction in popular culture, most notably in the ‘Jurassic Park’ and ‘Jurassic World’ films, has significantly shaped public perception of this dinosaur. The portrayal of Velociraptor as a large, intelligent, pack-hunting predator in the 1993 film, although scientifically inaccurate in size and behaviour, captured the imagination of millions and solidified its status as a cultural icon. This image, however, contrasts with the more accurate scientific reconstructions that depict Velociraptor as a smaller, feathered theropod. The gap between scientific understanding and public perception highlights the challenges and opportunities in science communication. While the feathered, bird-like depiction of Velociraptor has gradually gained acceptance, popular media has been slow to update its portrayal. Documentaries and educational programs incorporating the latest scientific findings have played a crucial role in bridging this gap, fostering a more accurate and nuanced understanding of Velociraptor among the general public.
As we look to the future, ongoing research and technological advancements promise to further unravel the mysteries of Velociraptor life and environment. Future discoveries, particularly in the areas of soft tissue preservation, isotopic analysis, ichnology, and biomechanical modelling, will likely provide new insights about the type of integumentary structures covering its body, its colour, reproduction (eggs), ontogeny, and the sound it was making. Moreover, continued exploration of the Gobi Desert and other Late Cretaceous sites may uncover additional specimens that can fill in gaps in our knowledge of Velociraptor and its closer taxa.
In summary, the centenary of the first description of Velociraptor is not just a celebration of a remarkable palaeontological find but also a testament to the dynamic nature of scientific inquiry. From its initial discovery to its status as a cultural icon, Velociraptor has played a pivotal role in our understanding of theropod evolution and the broader narrative of dinosaur research.
ELECTRONIC SUPPLEMENTARY MATERIAL
This article contains electronic supplementary material which is available to authorised users. In particular, a review of the systematic palaeontology of Velociraptorinae is provided as Appendix 1, and the consensus trees length and a diagnostic for each character in the conducted phylogenetic analyses are available as Appendix 2 and Appendix 3.